Evoluutio (kirja)
Evoluutio1 on maineikkaan evoluutiobiologi Ernst Mayrin kirjoittama evoluutioteorian yleistajuinen oppikirja.
Mitä evoluutio on[muokkaa]
Millaisessa maailmassa elämme?[muokkaa]
Hyvin pian kirjan alkuun Mayr esittelee kolme maailmanselitysmallia:
- Ikuinen maailma. Tämä oli kreikkalaisen filosofin Aristoteleen näkemys. Hän uskoi maailman olleen aina olemassa. Joidenkin filosofien mukaan maailma oli ikuisen lisäksi muuttumaton, toisten mielestä se kävi läpi erilaisia vaiheita. Näiden vaiheiden jälkeen maailma palasi taas varhaisemmalle asteelleen. Näkemys ei ole koskaan ollut suosittu, sillä ihmiset ovat tunteneet enemmän vetoa ajatukseen, että maailmalla olisi alku.2
- Nuori, muuttumaton maailma. Mayr väittää tämän olevan Raamattuun perustuva näkemys ja kertoo sen olleen vallassa keskiajalta 1800-luvun puoliväliin asti. Hän väittää, että kreationistit uskovat Jumalan luoneen kaikki eliölajit jo täysin sopeutuneiksi nykyisiin ekologisiin lokeroihinsa ja luomakunnan olevan samassa tilanteessa kuin luomisviikollakin.3 Nykyään edes nuoren maan kreationistit eivät usko näin. Kreationistit hyväksyvät eliöiden muuntelevan Jumalan luoman luodun tyypin4 sisällä. He uskovat maailman rapistuvan syntiinlankeemuksen takia, eivätkä pidä ekosysteemiä muuttumattomana, vaan uskovat sen mullistuneen esimerkiksi vedenpaisumuksen yhteydessä katastrofaalisesti.
Muuttumattoman maailman esitteleminen ainoana vanhan maailman vaihtoehtona on harhaanjohtavaa. Lajien tai koko ekosysteemin muuntumiskyvystä ei tietenkään voida vielä tehdä minkäänlaisia johtopäätöksiä koskien maan ikää. Mayr syyllistyy väärään vastakkainasetteluun, koska kyseessä on vain yksi Raamattuun uskovien kreationistien historian varrella esittämistä näkemyksistä. Nykyään kaikki merkittävimmät nuoreen luomiseen sitoutuvat kreationistijärjestöt irtisanoutuvat tästä näkemyksestä.5 - Kehittyvä maailma. Mayrin mukaan 1600-luvun tieteellisen vallankumouksen myötä päädyttiin siihen, että havainnot olivat ristiriidassa Raamatun kanssa. Pääteltiin maapallon huima ikä ja todettiin, ettei luonto ollutkaan staattinen. Mayr kertoo kuinka Raamatun ja luonnontieteellisten havaintojen ristiriidan selvittämiseksi kehiteltiin Scala Naturae eli suuri olemassaolon ketju. Mayr kirjoittaa evoluution olevan epäsäännöllinen prosessi, kuten esimerkiksi mannerlaattojen liikkeet. Mayr määrittelee evoluution seuraavasti:
Evoluutio on eliöiden muodostamien populaatioiden ominaisuuksissa ajan mittaan tapahtuvaa muutosta.
– Ernst Mayr, Evoluutio s.30
Mayrin mukaan kopernikaaninen vallankumous näytti ensimmäisenä toteen, että kaikkia Raamatun väitteitä ei voitu ottaa kirjaimellisena totuutena.6 Kopernikaaninen vallankumous tarkoittaa siirtymistä maakeskeisestä maailmankuvasta aurinkokeskeiseen. Kyseessä oli Ptolemaioksen 100-luvulla kehittämä maakeskeinen maailmankuva, jota tiedemaailma tuki ja kehitteli tuhannen vuoden ajan asettamatta sitä lainkaan kyseenalaiseksi. Myös katolinen kirkko tuki tätä näkemystä, vaikka Raamattu ei sitä edellytäkään.7
Evoluutio ja entropia[muokkaa]
Mayr ottaa esille myös toisinaan esitetyn väitteen siitä, että evoluutioteoria on ristiriidassa entropian lain kanssa, koska entropian lain mukaan pitäisi ajan kuluessa epäjärjestyksen lisääntyä, mutta evoluutioteorian mukaan järjestys lisääntyy. Hän selittää tämän sillä, että entropian laki toimii vain suljetuissa järjestelmissä, mutta evoluutio tapahtuu avoimessa järjestelmässä. Tämä taas tarkoittaa hänen mukaansa, että organismit voivat vähentää entropiaa ympäristön kustannuksella järjestelmässä, jossa aurinko tarjoaa koko ajan lisää energiaa.8 Väite ei ole täysin ongelmaton, sillä pelkkä auringon säteilemä energia ei johda kehitykseen. Hirsipinoon vuosikausiakaan paistava aurinko ei rakenna taloa, vaan pikemminkin hirsipino lahoaa vähitellen luonnollisten prosessien vaikutuksesta. Bensiininkin palamisesta saadaan enemmän hyötyä irti, kun se poltetaan monimutkaisessa moottorissa, jolloin bensiinin kemiallinen energia voidaan ohjata mekaaniseksi liike-energiaksi.9 Argumentilla voidaan kuitenkin osoittaa, ettei entropian laki estä paikallisen järjestyksen lisääntymistä, vaikka tekeekin ohjaamattomasta järjestyksen lisääntymisestä suuressa mittakaavassa epätodennäköistä.
Auringonvalon säteilyenergia ei esimerkiksi suoraan elätä ihmistä, vaan tämä energia täytyy ensin sitoa kemialliseen muotoon kasvien viherhiukkasissa. Tämän jälkeen säilötty energia täytyy ohjata niin, että se saadaan valjastettua elintoimintojen kannalta tarkoituksenmukaiseen käyttöön.10 Auringonvalon energia ei voi myöskään kasvattaa kasvia ilman solujen tarkoituksenmukaisesti toimivia molekyylikoneita, jotka ohjaavat energian kasvun kannalta tarkoituksenmukaisiin kemiallisiin reaktioihin.
Perinnöllisen muuntelun kohdalla ainoa ohjaava tekijä on kuitenkin luonnonvalinta, joka ei pyri kehittämään eliöitä monimutkaisemmiksi. Luonnonvalinnan kautta lajit muuntuvat ainoastaan sellaisiksi, että ne selviävät ja tuottavat mahdollisimman paljon jälkeläisiä mahdollisimman tehokkaasti vallitsevissa olosuhteissa. Tällainen muuntelu tapahtuu kuitenkin pääsääntöisesti tuhoamalla olemassaolevia monimutkaisia rakenteita, eikä rakentamalla uusia. Siksi auringon valon energia ei itsessään riitä rakentamaan eliöissä uudenlaista monimutkaisuutta.
Leedsin yliopiston termodynamiikan ja palamisen teorian professori Andrew McIntoshin mukaan termodynamiikka estää eliöiden geneettisen informaation lisääntymisen evoluutiossa. Hän pyrkii osoittamaan tutkimuksessaan, ettei maan kaltaisessa järjestelmässä auringosta saatava vapaa energia ratkaise tätä ongelmaa.11 Älykkään suunnittelun kannattaja Granville Sewell on myös esittänyt, että elämän monimutkaisten rakenteiden kehittyminen edustaa entropian vähenemistä, mikä rikkoisi termodynamiikan toista pääsääntöä ja sitä vastoin tukisi älykästä suunnittelua.12
- Granwille Sewell: A Mathematican's view of Evolution (video, 15 min)
Evoluution todisteita[muokkaa]
Seuraavaksi Mayr lähtee käsittelemään evoluutioteorian todisteita. Hänen mukaansa evoluutioteoria on varma totuus, jota tukevat kaikki todisteet biologian eri aloilta. Hänen mukaansa niitä ei voi itse asiassa edes ymmärtää ilman evoluutioteoriaa. Mayr argumentoi evoluutioteorian vakuuttavimman näytön olevan fossiileissa, joskin harmittelee fossiiliaineiston "töksähtelevyyttä". Tämän hän kuitenkin selittää sillä, että monet fossiilit ovat pyyhkiytyneet pois monissa luonnonmullistumissa. Lisäksi on hyvin epätodennäköistä, että eliö ylipäänsä fossiloituu, sillä se vaatii eliön hautautumisen nopeasti täysin hapettomaan tilaan13.
Archaeopteryx[muokkaa]
Silloin tällöin kuitenkin löytyy Mayrin mukaan onnekkaasti joku harvinainen välimuotofossiili, ja näistä esimerkiksi hän nostaa Archaeopteryxin. Hän kirjoittaa Archaeopteryxin olevan alkukantaisen linnun fossiili jurakaudelta, jolla oli monia esimuodoilta polveutuneita ominaisuuksia, kuten hampaat ja pitkä pyrstö. Lisäksi sillä oli monia lintumaisia piirteitä, kuten silmät, höyhenet, siivet ja aivot14.
Archaeopteryxin ylimenomuotouteen liittyy kuitenkin monia ongelmia. Kunnollisella ylimenomuodolla tulisi olla rakenteita, jotka ovat muuntumassa esi-isälajin rakenteesta lapsenlapsilajin rakenteeksi, mutta eivät edusta kumpaakaan. Sen sijaan Archaeopteryxillä on mosaiikkimaisesti sekä matelijoiden, että lintujen rakenteita. Tyypillisiä matelijan tuntomerkkejä ovat hampailla varustetut leuat, puuttuva rintalastan harja, kynnelliset raajat, selkänikamat, koukkulisäkkeettömät kylkiluut ja pitkä pyrstöranka. Linnuille tyypillisiä piirteitä puolestaan ovat höyhenpeite, eturaajan luusto sekä lantion ja jalkojen luusto. Esimerkiksi sulat eivät edusta välimuotoa matelijan suomujen ja linnun sulkien välillä, vaan ne ovat kaikin puolin samanlaiset kuin linnun sulat. Lisäksi matelijamaiset piirteet eivät ole yhteisiä millekään tietylle matelijaryhmälle, joten archaeopteryxin esi-isän määrittäminen on ongelmallista. Hampaat eivät myöskään ole itsestäänselvästi merkki matelijamaisuudesta, sillä kaikissa selkärankaisryhmissä on sekä hampaallisia että hampaattomia muotoja, ja näin on myös linnuissa. Archaeopteryxin hampaat muistuttavat krokotiilien hampaita, mutta krokotiilin kaltaisilla matelijoilla on erilainen lantio ja niiltä puuttuvat solisluut. Millään nykyisin tunnetulla linnulla ei ole samanlaisia nikamia kuin Archaeopteryxillä, mutta sen sijaan kalaliskoilla on vastaavat nikamat. Kalaliskot eivät kuitenkaan muiden rakennetuntomerkkiensä osalta sovellu esi-isäksi. Yleensä Archaeopteryxin esi-isänä pidetään Ornitischioita, lintulantioista dinosauruslahkoa, joilla ei kuitenkaan ole solisluita. Eturaajojen kynnet muistuttavat Saurichioiden eli liskonlantioisten dinosaurusten kynsiä, mutta niiden lantio on kuitenkin väärän mallinen. Toisaalta myös joillakin linnuilla kuten hoatsinilla ja turakolla on saman tyyppiset liikuteltavat kynnet. Archaeopteryx on siis tyypillinen mosaiikkimuoto, eikä sitä voida pitää selkeänä yhdistävänä linkkinä minkään kahden eliöryhmän välillä.15
Virallinen iänmääritys sijoittaa Archaeopteryxin sukupuussa "väärään kohtaan". Archaeopteryx on ajoitettu noin 153 miljoonaa vuotta vanhaksi, ja jopa nokkalintu Confuciusornis on ajoitettu 135 miljonaa vuotta vanhaksi. Polveutumisopin mukaiset Archaeopteryxin dinosaurusesi-isät, kuten Sinosauropteryx ja Caudipteryx on ajoitettu 125 miljoonaa vuotta vanhoiksi. Näin ollen Archaeopteryx ja Confuciusornis on ajoitettu miljoonia vuosia vanhemmiksi kuin niiden oletetut dinosaurus esi-isät. Lintutieteilijä Alan Feduccia onkin kritisoinut dinosauruksista lintuun -opinkappaletta toteamalla, että eihän kukaan voi olla isoisäänsä vanhempi. Dinosauruksista lintuihin -kehityskulkuun uskovat ovat toisinaan vastanneet tähän toteamalla, että isoisä voi elää pidempään, kuin lapsenlapsi. Jos tämä on totta, on vaikea ymmärtää, että kehittynyt nokkalintu Confuciusornis voisi ilmaantua 10 miljoonaa vuotta aikaisemmin, kuin sen höyhenpeitteiset esi-isät.16 Tämä edellyttää käytännössä sitä, että kaikki lapsenlapsilajia varhaisemmat isoisälajin edustajat ovat kadonneet fossiiliaineistosta.
Archaeopteryxillä on myös ainutlaatuisesti suunnitellut läpivirtauskeuhkot. Ilma kulkee keuhkoissa vain yhteen suuntaan mahdollistaen näin maksimaalisen hapenottokyvyn. Tämä järjestelmä on täysin erilainen, kuin matelijoiden paljekeuhkot, jollaiset ovat jopa Sinosauropteryxillä. Ensimmäisillä välimuodoilla olisi reikä palleassa17, jolloin luonnonvalinta toimisi sitä vastaan. Sen keuhkot vaativat myös tuekseen kiinnityksen reisiluihin, jotka dinosauruksilla ovat liikkuvat, eivätkä siten voi tarjota tukea lintumaiselle keuhkojärjestelmälle.18
Nisäkkäiden evoluutio[muokkaa]
Varhaisimpina nykyisin elävien nisäkkäiden eli synapsidien19 edustajina pidetään pelycosaureja20, jotka ovat evolutionistisessa luokittelussa lähempänä nisäkkäitä, kuin muita vesikalvollisia.19
Mayr mainitsee joidenkin fossiililinjojen olevan esimerkillisen täydellisiä. Esimerkeikseen hän ottaa kehityslinjan nisäkäsliskoista (Therapsidae) nisäkkäisiin, joiden kohdalla hän mainitsee joidenkin fossiilien olevan niin selviä välimuotoja, että niiden sanominen nisäkkääksi tai matelijaksi on täysin mielivaltaista.
Tässä ketjussa on kuitenkin suuria aukkoja. Fossiileista voidaan nähdä luusto, joten niistä ei voida nähdä joitakin nisäkkäiden oleellisia tuntomerkkejä, kuten verenkiertojärjestelmän ja munuaisten toimintaa, karvoitusta, maitorauhasia ja lämmönsäätelyn fysiologiaa. Näistä ominaisuuksista voidaankin esittää vain spekulaatioita. On myös huomattava, että luonnonvalinta voi valita vain koko organismin, ei sopivimpia geenejä tai yksitäisiä rakenteita.
Löydetyistä fossiileista ei voi mitenkään päätellä niiden välisiä geneettisiä sukulaisuuksia, ellei tällaisia jo ennalta oleteta olevan. Myös erikseen luotuihin perusryhmiin kuuluvat eliöt voivat fossiloitua, joten fossiilien löytyminen ei sinänsä todista polveutumisopin puolesta vaan se täytyy päinvastoin edellyttää oikeaksi, jotta fossiililöytöjen väliset polveutumissuhteet ylipäätään vaikuttaisivat mielekkäiltä tutkimuskohteilta. Kaikki riippuukin siis tulkinnasta, joka taas riippuu maailmankatsomuksellisista taustaolettamuksista.
Valaiden evoluutio[muokkaa]
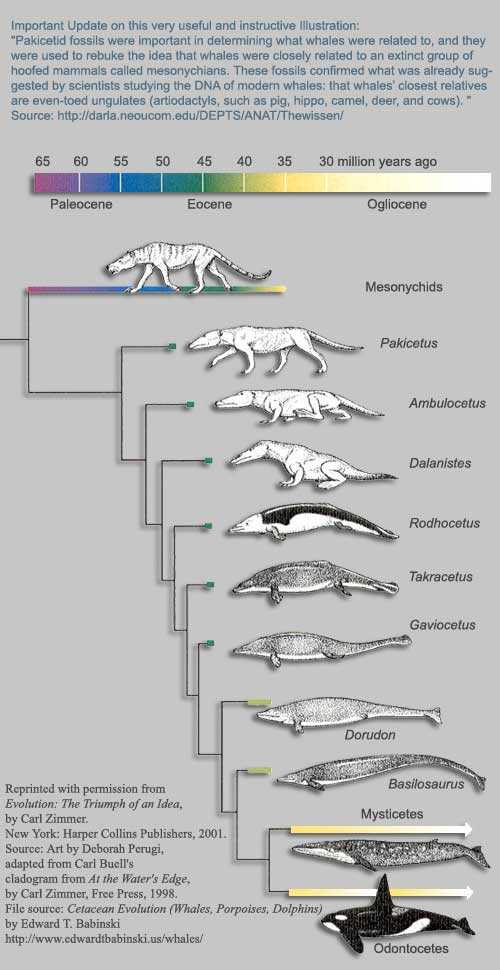
Mayr argumentoi, että erityisen täydellinen sarja fossiileja on löydetty myös valaiden esimuotojen (sorkka- ja kavioeläinten) ja nykyisten muotojen väliltä. Tämän päätelmän tueksi hän esittää kaavion välimuodoista.21
Mayrin esittämässä kaaviossa valaiden esi-isiksi esitellään Mesonychidit. Tätä näkemystä pidettiinkin ylivoimaisesti todistettuna ja sitä esiteltiin dogmaattisena totuutena myös esimerkiksi Yhdysvaltain kansallisen tiedeakatemian opettajien opaskirjassa.22 Nyt oletetut homologiset piirteet mesonychidien ja valaiden välillä23 on selitetty konvergenttisen evoluution tuloksiksi, eikä niillä olekaan enää mitään tekemistä yhteisen polveutumisen kanssa, kuten aiemmin väitettiin.24
Mayrin esittämä diagrammi näyttää hienolta, kun kaikki lajit on piirretty suunnilleen saman kokoisiksi. On kuitenkin huomattava, että esimerkiksi Basilosaurus on kymmenen kertaa pidempi kuin Ambulocetus.25
Mielessä on hyvä pitää myös, että Darwinilla ja hänen seuraajillaan ei pitkiin aikoihin ollut vähääkään uskottavan tuntuista fossiiliaineistoa valaiden evoluution puolesta, vaan valaiden kehityshistoria pysyi täysin uskonvaraisena evoluutioteorian haarana. Esimerkiksi valaiden evoluution asiantuntija E. J. Slijper myönsi vuonna 1962:
Meillä ei ole yhtäkään välimuotofossiilia edellä mainittujen maaeläinten (lihansyöjien ja sorkkaeläinten) ja valaiden välillä.26
Evoluutioteorialla oli näin ollen alusta asti suuri tarve pystyä esittämään tällaisia välimuotoja, joten ei ole mikään ihme, että ehdokkaita lopulta löydettiinkin – esimerkiksi Pakicetus27. Se on piirretty Mayrin kaaviossa ensimmäiseksi vedessä eläneeksi nisäkäslajiksi. Pakicetus hahmoteltiin alun perin muutamien löytyneiden luu- ja hammasfossiileiden ympärille. Kun sitten myöhemmin löydettiin loputkin kyseisen lajin luurangosta, se havaittiinkin vikkeläjalkaiseksi maaeläimeksi.28 Tämä on varsin tyypillinen esimerkki evolutionistien harhaanjohtavasta julkisuushakuisuudesta: liioiteltua "todistusaineistoa" rakennettiin evoluutioteoreettisen mielikuvituksen voimin muutaman luunpalasen varaan. Tarinan opetus onkin kiteytettävissä erään evolutionistin sanoin29:
Fossiilit ovat häilyväisiä. Luut laulavat sitä laulua, jota halutaan kuulla.
Toinen välimuotoehdokas, Basilosaurus30, oli käärmeen kaltainen, noin 21 metriä pitkä merinisäkäs, jolla oli 1,5-metrinen kallo: täysi vesieläin, jota on vaikea kuvitella maanisäkkäiden ja valaiden välimuodoksi. Selkärankaisia eliöitä tutkiva paleontologi ja evolutionisti Barbara Stahl huomauttaa:
Ruumiin käärmemäisyys ja poskihampaiden omalaatuinen muoto tekevät selväksi, että archaeocetes (kuten Basilosaurus) eivät voi olla modernien valaiden esi-isiä.31
On myös huomattava, että modernien valaiden – sekä hammasvalaiden (Odontoceti) että hetulavalaiden (Mysticeti) – rakennetyypit ilmestyvät fossiilikerrostumiin yhtäkkisesti32. Stahl huomauttaa myös, että kallon rakenne valaiden molemmissa rakennetyypeissä
ilmentää oudon muutoksen, johon ei ole pienintäkään viitettä Basilosauruksella eikä sen sukulaisilla: sieraimet ovat siirtyneet pään selkäpuolelle ja takaosaan, ja samaan aikaan nenän luut ovat pienenneet ja kulkeutuneet ylöspäin. Myös välileuan- ja leuanluut ovat laajenneet muodostaakseen suojan alkuperäiselle aivokopalle.31
Basilosauruksella tosin oli pienet takaraajat – varmasti liian pienet kävelyyn. Jotkut ovat siksi väittäneet niitä surkastumiksi. Eräät evolutionistitkin pitävät kuitenkin todennäköisempänä, että niitä on käytetty parittelun aikana parittelukumppanista kiinni pitämiseen. Esimerkiksi Philip Gingerich on todennut33:
Minusta tuntuu, että ne ovat voineet olla vain jonkinlainen seksuaali- ja lisääntymiselin.
Hevosen evoluutio[muokkaa]
Mayr pitää kaikkein parhaimpana todisteena evoluutiosta kuitenkin hevosten sukupuuta, jossa on hänen mukaansa täydellinen sarja fossiileita kantalajista (Eohippus) nykyiseen hevoseen (Equus).34
Hän esittää kuvitteellisen hevosen evoluution osoittavan sarjan. Nykyään monet evolutionistit itsekin myöntävät suoraan, että hevosen evoluution rakennelma on kärsinyt vararikon. Evolutionisti Boyce Rensberger sanoi – pitäessään puheen Chicagon luonnontieteen kenttämuseossa vuonna 1980 pidetyssä 150 evolutionistin nelipäiväisessä symposiumissa, joka käsitteli asteittaisen evoluutioteorian ongelmia – että hevosen evoluution rakennelmalla ei ole perusteita fossiiliaineistossa ja että ei ole havaintoja kehitysprosessista, joka vastaisi hevosen asteittaista evoluutiota.
Mayr esittää suositun hevosen evoluution esimerkin, jossa ehdotetaan asteittaisten muutosten sarjaa nelivarpaisesta, ketunkokoisesta 50 miljoonaa vuotta sitten eläneestä olennosta nykyiseksi yksivarpaiseksi ja paljon suuremmaksi hevoseksi. Tämän kehityslinjan on jo kauan tiedetty olevan väärä. Asteittaisen muuttumisen sijasta näiden välittävien lajien fossiilit viittaavat siihen, että kukin laji oli täysin erillinen, vallitsi muuttumattomana ja kuoli sitten sukupuuttoon. Siirtymämuotoja ei tunneta.
Englannin luonnonhistorian museon kuuluisa paleontologi Colin Patterson sanoi museon pohjakerroksessa yleisölle avoinna olleesta "hevosen evoluutiota" esittelevästä näyttelystä:
| ” | On kerrottu hurja määrä toinen toistaan mielikuvituksellisempia tarinoita siitä, millainen tämän (elämän) historian luonne todella on. Kuuluisa esimerkki on yhä alakerrassa esillä oleva, ehkä viisikymmentä vuotta sitten rakennettu hevosen evoluution esitys. Oppikirja toisensa jälkeen on esittänyt sen kirjaimellisena totuutena. Tämä on minusta todella valitettavaa, erityisesti koska tällaisia tarinoita tarjoavat ihmiset saattavat itse olla tietoisia siitä, että osa tästä aineksesta on luonteeltaan spekulatiivista. | ” |
Mikä siis on "hevosen evoluution" rakennelman perusta? Tämä rakennelma muodostettiin hämäävien kaavioiden avulla, jotka oli laadittu asettamalla sarjaksi fossiileja erillisistä, laajasti eri aikoina Intiassa, Etelä-Afrikassa, Pohjois-Amerikassa ja Euroopassa eläneistä lajeista yksinomaan evolutionistien rikkaan mielikuvituksen perusteella. Eri tutkijat ovat ehdottaneet yli 20 erilaista karttaa hevosen evoluutiosta. Evolutionistit eivät ole saavuttaneet yhteisymmärrystä sukupuista, jotka täysin eroavat toisistaan. Ainoa yhtymäkohta näissä rakennelmissa oli usko, että hevosen (equus) esimuoto oli eoseenikaudella, 55 miljoonaa vuotta sitten elänyt koiran kokoinen eläin eohippus. Mutta oletetut kehityslinjat eohippuksesta equusiin ovat täysin epäyhtenäiset.
Evolutionistinen tiedetoimittaja Gordon R. Taylor selittää tätä harvoin tunnustettua totuutta kirjassaan The Great Evolution Mystery ("Suuri evoluutioarvoitus"):
| ” | Mutta evoluutioteorian suurin heikkous on ehkä se, että paleontologit eivät ole onnistuneet löytämään vakuuttavia suvunkehityslinjoja tai eliöiden sarjoja, jotka edustaisivat olennaista kehityksellistä muutosta. Hevonen mainitaan usein ainoana täydellisesti viimeisteltynä esimerkkinä. Mutta tosiasia on, että linja eohippuksesta equusiin on hyvin säännötön. Sen väitetään osoittavan jatkuvaa koon kasvua, mutta jotkut muunnokset eivät todellisuudessa olleet suurempia kuin eohippus vaan sitä pienempiä. Näytteitä eri lähteistä voi yhdistää vakuuttavan näköiseksi sarjaksi, mutta ei ole mitään todisteita siitä, että ne todella muodostivat tällaisen sarjan ajassa. | ” |
Kaikki nämä seikat todistavat vahvasti, että Mayrinkin esittelemät hevosen evoluution kaaviot ovat vain mielikuvituksellisia ja epätodennäköisiä tarinoita.35
Fossiililinjojen luotettavuus[muokkaa]
Mayr argumentoi, että fossiiliaineistosta löydettävien kehityslinjojen luotettavuutta kasvattaa se, että fossiileja löytyy vain siitä kerrostumasta, mistä niitä oletetaankin löytyvän. Esimerkkinä hän mainitsee kirahvin kehittyneen 30 miljoonaa vuotta sitten ja mainitsee päätelmien ja laskelmien menevän pahasti sekaisin, jos kirahvi löydettäisiin 60 miljoonaa vuotta vanhasta kerrostumasta.36
Tokihan päätelmät menevät pahasti sekaisin, mutta se vaatii vain teorian laittamista uusiksi. Fossiileja kuitenkin löytyy evoluutioteorian näkökulmasta vääristä paikoista, kuten Archaeopteryxin, valaiden evoluution ja nisäkkäiden evoluution kohdalla on havaittu.
Fossiilien iänmääritys[muokkaa]
Mayr toteaa fossiilien iänmäärityksen olleen aiemmin arpomista mutta tilanteen olevan nykyisin parempi, sillä radiometriset iänmääritysmenetelmät tarjoavat tarkan keinon laskea vulkaanisista aineksista koostuvan kerrostuman ikä.37
Radiometrisetkin iänmääritysmenetelmät ovat lähinnä arpapeliä. Huomioitavaa on, että radiometrisillä iänmääritysmenetelmillä pyritään selvittämään aikaa, jolloin kivi on kivettynyt. Ei siis aineksen ikää, vaan kivettymishetkeä. Uudessa-Seelannissa yksi aktiivisimmista tulivuorista, nimeltään Ngauruhoe-vuori purkautui viimeksi vuonna 1975. Sen laavaa tutkittiin kalium-argon-menetelmällä, siten että näytteet otettiin silminnäkijätodistusten perusteella 25-51 vuotta vanhoista kerrostumista. Tutkimus tehtiin 2000-luvun alussa, jolloin kyseisen menetelmän virallinen mittausraja oli 270 000 vuotta. Jos menetelmä toimisi, se antaisi alle 270 000 vuotta vanhoille näytteille iäksi <270 000 vuotta. Iänmääritystä varten lähetettiin näytteitä hyvin arvostettuun kaupalliseen ajoituslaboratorioon38. Laboratoriolle ei kerrottu etukäteen, minkä ikäisiä näytteiden tiedettiin olevan. Neljä näytettä antoi kiviaineksen iäksi alle 270 000 vuotta, ja yksi alle 290 000 vuotta, mutta kahdeksalle näytteelle tuli jokin ikä väliltä 800 000 vuotta - 3 500 000 vuotta. Tämä tapaus on vain yksi monista vastaavista tapauksista, joissa radiometristen ikien antamat tulokset poikkeavat selvästi silminnäkijöiden havainnoista. Yksi ajoitusmenetelmien ongelma onkin, että niissä on lopputuotetta jo lähtötilassa, eikä nollahetkeä pystytä luotettavasti määrittämään.39
Luonnon monimuotoisuus ja eliöiden samankaltaiset ominaisuudet[muokkaa]
Mayr toteaa, että maailmassa näytti olevan vakava ristiriita: yhtäältä luonto oli käsittämättömän monimuotoista, toisaalta monilla eliöillä oli samankaltaisia ominaisuuksia. Esimerkiksi oli sammakoita, käärmeitä ja nisäkkäitä, jotka olivat anatomialtaan samantyyppisiä, mutta huomattavan erilaisia verrattuna vaikkapa hyönteisiin. Darwinin esittelemä yhteisen polveutumisen teoria selitti tämän ristiriidan sillä, että mikäli joillain lajeilla oli yhteneviä ominaisuuksia eroavaisuuksiensa lisäksi, ne on peritty kantamuodolta. Eroavaisuudet ovat kehittyneet vasta tämän jälkeen. 40
Täysin suvereenilla, kaikkivaltiaalla ja kaikkitietävällä Jumalalla on hyvin mahdollisuus luoda luonnosta käsittämättömän monimuotoista, mutta käyttää monilla eliöillä samankaltaisia ominaisuuksia. Tämä voikin johtua sopivien rakenneratkaisujen vähyydestä. Siten näiden kahden seikan välillä ei ole mitään ristiriitaa myöskään luomisnäkemyksen näkökulmasta katsottuna.
Polveutumisen tutkimus ja homologia[muokkaa]
Mayrin mukaan polveutumisen tutkimus on lähinnä homologisten ominaisuuksien tutkimista sen takia, että kaikki lajit ovat polveutuneet lähimmästä yhteisestä kantamuodostaan. Sen johdosta voidaan homologisten ominaisuuksien perusteella päätellä polveutuminen.41
Homologisiin päättelyihin liittyy kuitenkin kehäpäätelmä, joka selvitetään alla.
Mayr kirjoittaa homologian voivan hyvin onnistuneesti paikata evolutiivisten kehityslinjojen aukkoja. Tähän hän ottaa esimerkiksi Archaeopteryxin. Vähän myöhemmin hän lainaa homologian määritelmän Richard Owenilta, jonka mukaan homologia tarkoittaa erilaisten eläinten samaa elintä, jolla on kaikenlaisia käyttötarkoituksia ja muotoja. Kaksi elintä ovat sama elin silloin, kun evoluutio on johtanut ne kyseessä olevien lajien lähimmästä yhteisestä esimuodosta. Esimerkiksi tästä Mayr tarjoaa myyrän, apinan, valaan ja lepakon eturaajoja, jotka ovat muuntuneet hänen mukaansa kaivamiseen, kiipeilyyn, uimiseen ja lentämiseen. Raajan perusmuoto on Mayrin mukaan ollut jonkin kävelevän nisäkkään, esimerkiksi koiran, eturaaja. Väite siitä, että kahden aika etäistä sukua olevan lajiryhmän tietyt ominaisuudet olisivat homologisia, on aluksi vain arvaus. Päätelmää on testattava monilla eri kriteereillä, kuten ominaisuuden sijainnilla muihin elimiin verrattuna, kehittyykö ominaisuus samoin yksilönkehityksen aikana ja ominaisuuden sopusoinnussa oleminen muista homologioista saadun todistusaineiston kanssa. Homologia onkin aina pääteltävä, eikä sitä voida todistaa.42
Homologiset rakenteet eivät useinkaan synny samoista alkion osista, vaikka niin evoluutionämyksen perusteella niin ennustettiinkin.43 Itse asiassa on harvinaisuus, että ne syntyisivät samoista alkion osista.44 Niitä voidaan pitää paremminkin saman suunnittelijan toiminnan todisteena.45 Homologiaan liittyy myös seuraavanlainen, eräästä biologian oppikirjasta löytyvä kehäpäätelmä:
Homologisiksi rakenteiksi kutsutaan samantyyppisiä rakenteita, koska ne on peritty yhteiseltä esi-isältä.
Myöhemmin samalla sivulla lukee:
Yhdenmukainen rakenne on todiste yhteisen esi-isän olemassaolosta.46
Embryologia ja rekapitulaatio[muokkaa]
Käsitellessään embryologiaa Mayr käyttää vahvana evoluution todisteena Ernst Haeckelin sikiökuvia.47
Mayr kirjoittaa rekapitulaation tarkoittavan yksilön sikiökehityksen vaiheessa ilmestyviä ja sitten katoavia rakenteita. Esimerkiksi hetulavalaille kehittyy hampaat tietyssä sikiövaiheessa, mutta ne imeytyvät elimistöön ja katoavat myöhemmin. Mayr toteaa Haeckelin teorian "yksilönkehitys toistaa kehityshistorian" olleen liian pitkälle vedetty, mutta huomauttaa, että esimerkiksi kidustaskujen ilmaantuessa nisäkkäiden sikiöt toistavat alkumuodolla olleen tilanteen. Mayr argumentoi, etteivät tällaiset muutokset ole millään tavoin harvinaisia vaan että niitä tunnetaan tuhansittain eliökunnassa. Mayr selittää yksilönkehityksen menevän kiertotietä eikä suoraan, sen tähden että esimuodoilta perityt ominaisuudet toimivat sikiönkehityksen organisaatiokeskuksina, kun siirrytään seuraaviin kehitysvaiheisiin. Esimerkkinä Mayr käyttää havaintoa, että kun sammakkoeläimen sikiöltä katkaistiin alkumunuaisen johdin, ei esimunuainen kehittynyt. Nisäkkäiden kiduksia koskien Mayr jatkaa, että kidusmaisia rakenteita ei käytetä koskaan hengittämiseen, vaan niistä tulee monet nisäkkäiden niskan seudun rakenteet, kun ne myöhemmässä yksilön kehityksen vaiheessa järjestellään uudelleen. Mayrin mukaan rekapitulaatio koskeekin vain eliöiden yksittäisiä rakenteita, ei koskaan esimuodon täydellistä aikuisvaihetta.48
Mayr spekuloikin hyvin vakuuttavan tuntuisesti lajien välisistä suhteista eri eliöiden sikiöissä ja myöhemmissä kehitysvaiheissa olevien samankaltaisuuksien perusteella. Tällaiset spekulaatiot ovat kuitenkin edelleen todistamattomia hypoteeseja, ja niiden tueksi esitettyjä havaintoja tulkitaan lähtöoletuksista riippuen eri tavoin: samat havainnot voidaan yhtä hyvin selittää luomisnäkökulmasta. Esimerkiksi ihmisen ja selkärankaisten alkionkehityksessä havaittavat yhtäläisyydet selittyvät sillä, että jokainen ihmisalkion kehitysvaihe on välttämätön seuraavalle vaiheelle ja koko ihmisen rakentumiselle. Kehitysvaiheet voidaankin näin ymmärtää ilman evoluutiota. Ihminen on ihminen alusta asti.49 Aihetta käsitelty myös artikkelissa: Rekapitulaatioteoria
Surkastumat[muokkaa]
Mayr mainitsee monilla eliöillä olevan osittain tai täysin toimimattomia rakenteita. Esimerkeikseen hän ottaa ihmisen umpisuolen, hetulavalaiden sikiöiden hampaat ja luolaeläinten silmät; hänen mukaansa ne valaisevat evoluution aikaisempaa kulkua. Hän argumentoi näiden edellä mainitsemiensa kolmen todisteen – surkastuneiden elimien, samankaltaisten sikiöiden ja rekapitulaation – asettavan ylitsepääsemättömiä ongelmia kreationistisen selityksen tielle mutta olevan sopusoinnussa yhteisen polveutumisen teorian kanssa.50
Surkastuneet elimet eivät kuitenkaan suoranaisesti kelpaa korkeammalle kehittymisen todisteiksi. Silloin jos elimellä ei ole mitään toimintoa, tai kun toiminto on rakenteeseen nähden liian vähäinen, on yleensä perusteltua olettaa että eliö on polveutunut sellaisesta kantamuodosta, jolla kyseinen elin on ollut toimiva. Silloinkin surkastuneet elimet todistavat enintään sen, että elimet voivat surkastua eli menettää toimintakykyään.51
Mayrin mainitsemat surkastumaesimerkit ovat sitä paitsi ongelmallisia, sillä ihmisen umpilisäkkeellä (kansanomaisesti umpisuolella) on olemassa rakenteeseen nähden täysin riittävä funktio: se toimii erityisesti ensimmäisten elinvuosien aikana taudinaiheuttajien torjunnassa niin sanottuna paksusuolirisana.52 Lisäksi vuonna 2007 julkaistun tutkimuksen mukaan umpilisäkkeellä on tärkeä tehtävä myös aikuisilla, sillä se säilöö suolistobakteereita ihmisen sairastuessa esimerkiksi koleran tai ameeban aiheuttamaan ripuliin. Ihmisen parannuttua umpilisäkkeessä säilynyt bakteerikanta synnyttää uudestaan kaikki elimistölle tarpeelliset bakteerit. Kehitysmaissa ripuli on yksi yleisimpiä kuolinsyitä.53
Hetulavalaiden sikiöiden hampaita koskien evolutionistit väittävät niiden todistavan hetulavalaiden kehittyneen hammasvalaista, mutta he eivät ole esittäneet mekanismia, joka romuttaisi täysin toimivan järjestelmän (eli hampaat) ja loisi tilalle aivan erilaisen järjestelmän (hetulat). Sikiöiden hampaiden funktioina on myös toimia eräänlaisina oppaina, jotta leukaperät muodostuisivat oikein.54
Luolaeläinten silmät taas kuuluvat taantuvan mikroevoluution piiriin. Esimerkiksi sokeiden luolakalojen silmät ovat surkastuneet, mutta valoisissa lähivesissä elää niiden näkökykyisiä lajitovereita. Sokeat luolakalat pystyvät lisääntymään näkevien lajitovereittensa kanssa ja saamaan lisääntymiskykyisiä jälkeläisiä. Luolassa, jossa on täysin pimeää, silmien menettämisestä on hyötyä, sillä silloin otaksuttavasti ravinteita säästyy ja silmien tulehdusvaara poistuu.55 Kyseessä on siten lajin sisäinen taantuma, jota ei voi käyttää todisteena korkeammalle kehittymisestä. Se toimii vain todisteena Mayrin esittelemää vanhentunutta kreationistista käsitystä vastaan.
Nämä kolme viimeksi mainittua todistetta eivät – Mayrin toteamuksesta huolimatta – aseta ylitsepääsemättömiä ongelmia kreationistisen selityksen tielle; ne asettavat niitä vain Mayrin esittelemälle vanhentuneelle luomiskäsitykselle. Surkastuneet elimet ovat sopusoinnussa luomisnäkemyksen kanssa, samankaltaiset sikiöt ovat sata vuotta vanha väärennös, ja rekapitulaatioksi tulkitut yksilönkehitysilmiötkin voidaan ongelmitta selittää luomisnäkemyksellä. Kuten evolutionisti Scadding on sanonut, "... surkastuneet elimet eivät ole evoluutioteorian todisteita." 56
Biomaantiede[muokkaa]
Mayr argumentoi, että näennäisen tempoilevia levinneisyyskuvioita ei voi selittää helposti luomisen tuloksena. Esimerkkeinä näistä epäsäännöllisistä kuvioista hän esittää Pohjois-Amerikan ja Euroopan eläimistöjen suhteellisen samankaltaisuuden verrattuna Afrikan ja Etelä-Amerikan faunojen huomattaviin eroihin, samoin Australian eläimistön omalaatuisuuden ja sen, ettei valtamerien saarilla yleensä ole nisäkkäitä. Hän kertoo evoluutioteorian auttaneen selittämään nämä ilmiöt.57
Mayr jatkaa jo ensimmäisessä luvussaan esittelemäänsä virheellistä vastakkainasettelua. Näennäisen tempoilevia levinneisyyskuvioita ei ole helppo selittää muuttumattoman maailman näkökulmasta, mutta nykyisen vallalla olevan kreationistisen käsityksen mukaan levinneisyyskuviot ovat hyvin selitettävissä.
Nykyään on myös löydetty monia evoluutioteorian näkökulmasta hankalasti tulkittavia havaintoja. Tällainen havainto on monien lajien tai kokonaisten heimojen esiintyminen valtavien välimatkojen päässä toisistaan. Esimerkiksi Siniharakoita asuu Kiinassa ja eräällä Espanjan kulmalla. Eräs salamanterilaji (Hydromantes) asustaa hyvin rajatuilla alueilla läntisen Välimeren alueella ja Kaliforniassa. Muurahaiskarhuja taas asustaa vain Etelä-Amerikassa nykyään, mutta fossiileina niitä esiintyy myös Saksassa. On myös huomattavaa, että ryhmä lähisukuisia makean veden kaloja esiintyy eri maanosissa. Koska kalat ovat sopeutuneet makeaan veteen, ne eivät voi ylittää valtameriä.58
Molekyylibiologia[muokkaa]
Mayr pitää yhtenä molekyylibiologian mukavimmista löydöistä sitä, että ruumiin makrorakenteiden ja molekyylitason muutokset liittyvät kiinteästi toisiinsa. Siten niissä tapauksissa, joissa morfologinen eli ulkonäöllinen tutkimus oli tuottanut suuria epäilyksiä eliöiden sukulaisuussuhteista, voitiin molekyylitutkimusten avulla paljastaa eliöiden todellinen suhde.59
Molekyylivertailujen tekeminen eliöiden sukulaisuussuhteita vertaillessa tuottaa kuitenkin monia ongelmia evoluutioteorian kannalta. Esimerkiksi aasin maito on lähes täysin samanlaista äidin maidon kanssa. Toistaan hyvin kaukana olevista taksonomisista ja erilaisissa kasviryhmissä esiintyy kofeiinia ja nikotiinia. Jos taas vertaillaan insuliineja, on kalkkarokäärme läheisempää sukua ihmiselle, kuin marsu. Riippuen siitä, mitä verrataan, on ihminen joskus läheisempää sukua banaanille, kuin simpanssille.60
Molekyylikello[muokkaa]
Mayr toteaa, että fossiiliaineiston puutteellisuuden vuoksi monien evolutiivisten kehityslinjojen geologisesta iästä oli mahdotonta saada tietoa. Hän mainitsee Zuckerkandlin ja Paulingin vuonna 1962 osoittaneen, että monet molekyylit muuttuvat aikojen saatossa hyvin tasaisesti. Nämä molekyylit voivat siten toimia molekyylikellona. Kukin molekyylikello voidaan ajastaa mittaamaan oikeaa aikaa nykyisten jälkeläisten tarkasti ajoitettujen fossiilien avulla. Mayr kuitenkin toteaa, että kyseistä menetelmää on käytettävä varovasti, sillä molekyylikellot eivät käy lainkaan niin tasaisesti kuin yleisesti ajatellaan. Sen lisäksi, että eri molekyyleillä on erilainen muutosvauhti, myös yksittäiset molekyylit voivat muuttaa muutosvauhtiaan, kun aikaa kuluu. Tätä kutsutaan mosaiikkievoluutioksi. Mayrin mukaan ristiriitaisissa tilanteissa pitää joko etsiä toinen fossiili tai määrittää jonkun toisen molekyylin muutosvauhti.61
Mayrin mainitsemat ongelmat molekyylikelloissa ovat merkittäviä, sillä molekyylikello ei ole voinut käydä samaan tahtiin niin monien eri organismien kaikissa vastinproteiineissa.62 Tällaisessa tapauksessa useimpien organismien mutaationopeuksien olisi oltava keskenään yhtä suuret. Havainnot kuitenkin osoittavat, että mutaationopeudet riippuvat eliösukupolvien määrästä eivätkä kuluneesta ajasta sinänsä, joten mutaatioita tulisi kertyä paljon nopeammin niihin organismeihin, joiden sukupolvet vaihtuvat tiuhimpaan tahtiin, kuten bakteereihin, ja paljon hitaammin harvakseltaan lisääntyviin lajeihin, kuten esimerkiksi norsuihin. Hyönteissukupolvetkin ovat sangen vaihtelevan mittaisia – ne kestävät kärpästen muutamista viikoista kaskaiden useisiin vuosiin – eivätkä vaikkapa kärpäset ole silti etääntyneet oletetusta yhteisestä kantamuodosta kaskaita pitemmälle. Niinpä teoria, jonka mukaan molekyylivertailujen säännönmukaisuudet olisivat parhaiten selitettävissä elämän hitaan kehityksen myötä vähitellen kertyneillä mutaatioilla, ei ole havaintoaineiston vahvistama.63
Uudet geenit[muokkaa]
Mayr argumentoi uusien geenien syntyvän pääosin jo olemassaolevan geenin kahdentuessa ja liittyessä kromosomiin edeltäjänsä viereen. Ajan myötä uusi geeni voi saada uuden tehtävän mutta vanha geeni jää edelleen toimittamaan vanhaa tehtäväänsä. Mayrin mukaan kahdentuminen ei aina koske vain yksittäistä geeniä vaan voi koskea myös koko kromosomistoa tai genomia.64
Mutaatioita ei kuitenkaan esiinny ainoastaan geenien kaksoiskappaleissa; niitä esiintyy läpi genomin. Siitä syystä kaikkien muiden genomin sisältämien vahingollisten mutaatioiden täytyy tulla eliminoiduiksi kelvottomien kuoleman kautta. Selektiiviset mutaatiot tavoitteena olevassa duplikaattigeenissä ovat äärimmäisen harvinaisia – eläimen genomissa se saattaa edustaa ainoastaan 1 osaa 30 000:sta. Mitä isompi genomi, sitä suurempi ongelma, sillä mitä suurempi genomi, niin sitä alhaisempi on mutaatioiden määrä, jonka eläin voi kestää ilman virhekatastrofia; tuloksena tästä, minkä tahansa duplikaattigeenin mutaation ilmaantuminen vie enemmän aikaa, puhumattakaan toivotunkaltaisesta mutaatiosta. Aikaa ei ole yksinkertaisesti ollut riittävästi jotta kyseisen kaltainen naturalistinen prosessi voisi selittää näkemiemme elävien olentojen sisältämän geneettisen informaation.65
Kokonaisen kromosomin duplikaatio on yleensä haitallinen, kuten on esimerkiksi Downin syndrooman yhteydessä. Lisäykset ovat lisäksi erittäin tehokas tapa tuhota totaalisesti olemassa olevien geenien toimivuus. Biofyysikko, tohtori Lee Spetner analysoi kirjassaan Not By Chance esimerkkejä mutaatioiden aiheuttamista muutoksista, joiden evolutionistit ovat väittäneet olleen informaatiota lisääviä muutoksia, ja osoittaa, että ne ovat itse asiassa olleet esimerkkejä spesifioitumisen menetyksestä, joka tarkoittaa, että niissä on ollut kyse informaation menetyksestä66.65
Johtopäätökset[muokkaa]
Johtopäätöksissään Mayr kertoo, että mitä tahansa biologian aluetta tutkitaan, se näyttää vastaansanomatonta todistusaineistoa evoluution puolesta. Hänen mukaansa tässä luvussa esitetyille tosiasioille ei ole muuta luonnollista selitystä kuin evoluutio.67
Ylläolevan kritiikin valossa Mayrin väite siitä, että mitä tahansa biologian aluetta tutkitaankin, se tuottaa vastaansanomatonta todistusaineistoa evoluution puolesta, on hyvin kyseenalainen.
Elämän synty ja sen alkuvaiheet[muokkaa]
Myöhemmin Mayr esittelee evoluutioteoreettista näkemystä elämän synnystä aina lintujen syntyyn asti historiallisena totena. Hän aloittaa tämän osan kirjastaan kertomalla, että maa syntyi noin 4,6 miljardia vuotta sitten geofyysisten ja tähtitietellisten todisteiden mukaan. Hänen mukaansa elämä kehittyi pian sen jälkeen, kun se tuli mahdolliseksi, eli noin 3,8 miljardia vuotta sitten. Elämä alkoi yhteen kerääntyneistä makromolekyyleistä, jotka kykenivät ottamaan energiaa ja ainetta auringon säteilystä ja elottomista molekyyleistä.68
Elämän syntyyn liittyy hyvin monia ongelmia, joita on käsitelty Abiogeneesi-artikkelissa. On myös hyvin kyseenalaista, että eräät bakteerit ja sinibakteerit olisivat pysyneet 3,5 miljardia vuotta samanlaisina, vaikka mutaatiot pommittavat niitä koko ajan.
Bakteerien morfologinen69 pysyvyys[muokkaa]
Hän toteaa varhaisimpien eliöfossiilien näyttävän hyvin samanlaisilta kuin jotkin nykyiset bakteerit ja sinibakteerit.70 Hänen mukaansa sinibakteerien huomionarvoisin seikka onkin niiden morfologinen pysyvyys. Tätä hän selittää sillä, että sinibakteerit lisääntyvät suvuttomasti, elävät hyvin suurissa populaatioissa ja pystyvät selviytymään ääriolosuhteissa. Hänen mukaansa bakteerit hallitsivat 1000 miljoonaa vuotta maan historiasta.71
Bakteerit ovat hajottajia, joten voidaan perustellusti kysyä, söivätkö nämä bakteerit toisiaan, kun tuottajat eli aitotumaiset eliöt syntyivät vasta tuhat miljoonaa vuotta myöhemmin? Tämä hypoteesi toimii melko heikosti, sillä bakteerit söisivät heti alkuunsa toisensa sukupuuttoon. Joidenkin sinibakteerien kyky hyödyntää ilmakehän typpeä ei ratkaise ongelmaa.72
Alkuilmakehä ja elämänsyntyteorioiden kehitys[muokkaa]
Mayr argumentoi, että maapallon varhaisvaiheen ilmakehässä ei ollut happea, vaan se koostui mm. metaanista ja vesihöyrystä.73 Hänen kuvaamansa tilanne on kuitenkin mahdoton, sillä kaikkialla, missä on vettä, on ennemmin tai myöhemmin myös happea.74
Mayrin mukaan ensimmäiset vakavasti otettavat teoriat elämän synnystä kehiteltiin 1920-luvulla, tekijöinä hän manitsee Oparinin ja Haldanen. Hänen mukaansa elämän synnystä on laadittu kuusi tai seitsemän kilpailevaa teoriaa, joista mikään ei tosin ole tyydyttävä. Mayrin mukaan elämän synnyn ongelma ei kuitenkaan ole enää niin suuri kuin se oli 1900-luvun alussa.75
Mayr tunnustaa, että elämää ei ole kukaan pystynyt tuottamaan laboratoriossa, mutta toteaa heti perään, että ei se mahdotonta voi olla, koska elämä syntyi heti sen jälkeen, kun sen syntyminen tuli mahdolliseksi 3,8 miljardia vuotta sitten.76
Mitokondrioiden ja viherhiukkasten alkuperä[muokkaa]
Mayr väittää mitokondrioiden olleen alun perin purppurabakteerien alfa-ryhmään kuuluneita bakteereja, kasvien viherhiukkasten puolestaan sinibakteereita.77 Mayrin ehdottama endosymbioositeoria on kuitenkin koottu hakemalla toisistaan irrallaan olevia samankaltaisuuksia, kuten dna:n rengasmaisuus, mutta kokonaisuutena teoria ei kestä kummassakaan tapauksessa.
Endosymbioositeorian mukaan mitokondrioiden tulisi olla huomattavan itsenäisiä yksiköitä solussa, mutta näin ei ole, sillä esimerkiksi ihmisen mitokondrion dna:lla koodataan vain 13 proteiinia, vaikka mitokondriot käyttävät toimintaansa ainakin 83 proteiinia.78 Mitokondrioista on tunnistettu 615 eri proteiinia, joten ne käyttävät luultavasti paljon muitakin proteiineja, mutta näiden kaikkien toimintaa ei tunneta.79 Lisäksi mitokondrio-dna:lla koodatuissa proteiineissa on laajoja alayksiköitä, jotka on koodattu solun dna:lla. Lisäksi rengasmaisuus on luonteva seuraus dna:n pienestä määrästä. Kun dna:a on paljon, se kannattaa hajauttaa useaan paikkaan informaation käsittelyn nopeuttamiseksi.
Mitokondrioiden säätelyjärjestelmä solussa on myös erittäin tarkka ja sen vioittumisen seuraukset ovat tuhoisia. Isompien solujen bakteerien nieleminen on myös hyvin yleistä ja sitä tapahtuu lukemattomia kertoja joka tunti. Kuitenkaan missään vaiheessa ei ole havaittu mitään mitokondrioiden syntyyn viittaavaa.78 Endosymbioositeoriaan ei ole mitään pakottavaa syytä uskoa, ellei oleta jo etukäteen, että polveutumisoppi on totta.
Kambrikauden räjähdys[muokkaa]
Mayrin mukaan kambrikauden räjähdys ei ole vakava ongelma, koska tuohon aikaan monet pehmeäruumiiset pääjaksot saivat tukirangan hyvin samanaikaisesti saalistuspaineen tai ilmakehän happipitoisuuden kasvun takia. Näin ollen aikaisemmista muodoista ei ole jäänyt fossiileja.80 Mayr tunnustaa, että fossiiliaineisto varhaisista kasveista on niukkaa. Hän kuitenkin toteaa, että morfologisten ja molekyylimenetelmien avulla on saatu koppisiemenisten kasvien polveutumissuhteista hyvä kuva.81
Selkärankaisten synty ja lintujen alkuperä[muokkaa]
Tämän aiheen käsittelyn aloittaessaan Mayr toteaa, että tapaamme suuressa luonnonhistoriallisessa museossa monia selkärankaisten alajaksoon kuuluvia lajeja. Mayrin mukaan nämä kehittyivät yksisoluisten eräästä ryhmästä, kaulussiimaeliöistä. Niistä kehittyivät simpukkaeläimet, joista taas kehittyivät diploblastiset onteloeläimet. Näistä puolestaan kehittyivät triploblastiset kaksikylkiset eläimet, jotka jakautuivat alku- ja jälkisuisiin, jotka viimemainitut jakautuvat edelleen neljään pääjaksoon. Mayr valittelee varhaisimpien selkärankaisten niukkaa fossiiliaineistoa mutta toteaa hiljattain löydetyn pari selkärankaista kalaa 530 ja 520 miljoonaa vuotta vanhoista kerrostumista.82 Hän käsittelee myös lintujen alkuperää ja jatkaa Archaeopteryxiä koskevia selvityksiään, joita on kritisoitu jo aiemmin tässä artikkelissa.83
Evoluutioteorian kannalta hyvin merkittäviin kysymyksiin kuuluu se, miten ulkoisesta tukirangasta siirryttiin sisäiseen tukirankaan. Sen ratkaisemiseksi tulisi selittää, miten luut kehittyivät hyönteisten kitiinipanssareita tai simpukan kuoria vastaavista rakenteista tai pehmeistä kudoksista. Tätä ei ole havaittavissa fossiiliaineistosta, eikä pätevää teoreettista malliakaan ole kehitetty. Luiden, nikamien, jänteiden ja niihin kuuluvien lihasten järjestelmän synty on myös hyvin monimutkainen tapahtuma, joten se vaatii huomattavasti aikaa, ja tästä tapahtumasarjasta pitäisi olla jäänyt useita välimuotoja. Ensimmäisiä kalojen jäänteitä on kuitenkin löydetty, kuten Mayrkin mainitsee, 530-520 miljoonaa vuotta vanhoista kerrostumista. Sisäiseen tukirankaan johtaneelle tapahtumasarjalle on siten melko vähän aikaa, eikä fossiileja ole löytynyt, vaikka esikambrikauden kerrostumat ovat täysin kykeneviä säilyttämään fossiilien hienorakenteen.84
Johtopäätökset[muokkaa]
Johtopäätöksissään Mayr toteaa, että kaikkein tyydyttävintä on se, että löydöt ovat sopusoinnussa Darwinin yhteisen polveutumisen teorian kanssa. Fossiiliaineisto yhdessä molekyylimenetelmien kanssa tuottaa kiistatonta todistusaineistoa evoluution puolesta. Varsieväkalojen ajateltiin aiemmin kuolleen sukupuuttoon, koska niitä tunnettiin vain 60 miljoonaa vuotta vanhoina fossiileina. Viimeisen 50 vuoden aikana on kuitenkin löytynyt kaksi elävää varsieväkalalajia. Tällaiset yllättävätkin löydöt ovat Mayrin mukaan aina sopineet ilman ongelmia darvinistiseen kehyskertomukseen.85
Viitteet[muokkaa]
- ^ Evoluutio. (What evolution is, 2001.) Suomentanut Jani Kaaro. Tieteen huiput. Helsinki: WSOY, 2003. ISBN 951-0-27897-1.
- ^ Evoluutio, s. 24
- ^ Kyseessä on William Paleyn kelloseppävertaus. Hänen mukaansa Jumala on kuin kelloseppä: hän on luonut lajit täydellisesti sopeutuneiksi eri olosuhteisiin niin, että ne eivät muutu.
- ^ Englanniksi "kind", eri merkitys kuin sanalla laji (engl. "species").
- ^ Katso esimerkiksi
* Answers in Genesis: "Species do change. Since Darwin’s day, many observations have confirmed this. In fact, new species have even been shown to arise within a single human lifetime -- Contrary to the accepted wisdom of Darwin’s day, the Bible nowhere teaches that species are fixed and unchanging. Rather, the book of Genesis refers to “kinds” -- and suggests that living things have had a very dynamic history. For example, as a consequence of the Fall, some animals became predators, and disease entered into the world. And after the Flood destroyed life on earth, God commanded the creatures on the Ark to “breed abundantly in the earth, and be fruitful, and multiply upon the earth”"
* Creation Ministries international: "Nowhere does Scripture teach the fixity of species,9 an erroneous belief that was held by several early biologists10 but which we know to be false today.11" - ^ Evoluutio, s. 26
- ^ Pekka Reinikainen: Unohdettu Genesis. Luominen vai evoluutio, s. 9. Kustannus Oy Uusi Tie, 1991. 2. täydennetty painos 1994. ISBN 951-619-239-4.
- ^ Evoluutio, s. 30-31
- ^ Pekka Reinikainen: Unohdettu Genesis. Luominen vai evoluutio, s. 66. Kustannus Oy Uusi Tie, 1991. 2. täydennetty painos 1994. ISBN 951-619-239-4.
- ^ Ihmisen täytyy ensin syödä näitä kemiallista energiaa sisältäviä molekyylejä, jotka ruuansulatuselimistö ohjaa verenkiertoon. Verenkierron kautta nämä ravintomolekyylit jaetaan ympäri elimistöä. Lopulta yksittäiset solut tuottavat näiden ravintomolekyylien kemiallisen energian avulla toisenlaisia molekyylejä, joissa energia on tehokkaammin käytettävissä elintoimintojen kannalta tarkoituksenmukaisella tavalla.
- ^ A. C. McIntosh, “Information and Entropy—Top-Down or Bottom-Up Development in Living Systems?” International Journal of Design & Nature and Ecodynamics 4(4) (2009): 351-385
- ^ Sewell, Granville: Evolution's Thermodynamic Failure Discovery Institute. Viitattu 6.7.2008.
- ^ Evoluutio s. 38-39
- ^ Evoluutio s. 39
- ^ Siegfried Scherer, Reinhard Junker: Evoluutio – kriittinen analyysi, s. 219-221. Datakirjat, 2007. ISBN 951-98558-0-7.
- ^ Jonathan Sarfati: The Greatest Hoax On Earth? Refuting Dawkins On Evolution, s. 124. Creation Book Publishers, 2010. ISBN 978-0-949906-73-1.
- ^ Pallea toimii niin sanottuna mäntänä matelijoiden paljekeuhkoissa
- ^ Jonathan Sarfati: The Greatest Hoax On Earth? Refuting Dawkins On Evolution, s. 125. Creation Book Publishers, 2010. ISBN 978-0-949906-73-1.
- > 19,0 19,1 http://en.wikipedia.org/w/index.php?title=Synapsid&oldid=553709357
- ^ http://en.wikipedia.org/w/index.php?title=Pelycosaur&oldid=551547766
- ^ Evoluutio s. 40, kaavio sivulla 42. Samanlainen kaavio on katsottavissa myös täällä, viitattu 11.5.2013
- ^ Teaching about Evolution and the Nature of Science (1998), viitannut Jonathan Sarfati: The Greatest Hoax On Earth? Refuting Dawkins On Evolution, s. 136. Creation Book Publishers, 2010. ISBN 978-0-949906-73-1.
- ^ erityisesti hampaiden ja kallon rakenteet
- ^ Jonathan Sarfati: The Greatest Hoax On Earth? Refuting Dawkins On Evolution, s. 136. Creation Book Publishers, 2010. ISBN 978-0-949906-73-1.
- ^ Jonathan Sarfati: The Greatest Hoax On Earth? Refuting Dawkins On Evolution, s. 137. Creation Book Publishers, 2010. ISBN 978-0-949906-73-1.
- ^ Slijper, E.J.: Dolphins and Whales, s. 17. University of Michigan Press, 1962. , viitannut Jonathan Sarfati: The Greatest Hoax On Earth? Refuting Dawkins On Evolution, s. 136. Creation Book Publishers, 2010. ISBN 978-0-949906-73-1.
- ^ ”valas Pakistanista” t. "Pakistanin valas"
- ^ Thewissen, J.G.M., ja muut, Skeletons of terrestrial cetaceans and the relationship of whales to artiodactyls, Nature 413:277-281, 20.9.2001, viitannut Jonathan Sarfati: The Greatest Hoax On Earth? Refuting Dawkins On Evolution, s. 138. Creation Book Publishers, 2010. ISBN 978-0-949906-73-1.
- ^ Shreeve, J., Argument over a woman, Discover 11(8):58, 1990 (viitattaessa ihmisen evoluutioon, mutta sopiva silti.), viitannut Jonathan Sarfati: The Greatest Hoax On Earth? Refuting Dawkins On Evolution, s. 138. Creation Book Publishers, 2010. ISBN 978-0-949906-73-1.
- ^ kreik. "liskokuningas" t. "kuningaslisko"
- > 31,0 31,1 Stahl B.J., Vertebrate History: Problems in Evolution, s. 489, Dover, 1985, viitannut Jonathan Sarfati: The Greatest Hoax On Earth? Refuting Dawkins On Evolution, s. 138-139. Creation Book Publishers, 2010. ISBN 978-0-949906-73-1.
- ^ valmiina ilman esimuotoja
- ^ Lainattuna The Press Enterprise -lehdessä 1.7.1990 s.A-15, viitannut Jonathan Sarfati: The Greatest Hoax On Earth? Refuting Dawkins On Evolution, s. 139. Creation Book Publishers, 2010. ISBN 978-0-949906-73-1.
- ^ Evoluutio s. 40, kaavio sivulla 43
- ^ http://www.netikka.net/mpeltonen/evopetos.htm Luettu 11.11.2010
- ^ Evoluutio s. 45
- ^ Evoluutio s. 40-45
- ^ Geochron Laboratories, Gambridge, Massachusetts
- ^ Marvin L. Lubenow: Myytti apinaihmisistä - kiista fossiilien ajoituksesta, s. 242-243. Datakirjat, 2005.
- ^ Evoluutio s. 49
- ^ Evoluutio s. 40
- ^ Evoluutio s. 54-55
- ^ Siegfried Scherer, Reinhard Junker: Evoluutio – kriittinen analyysi, s. 157-158. Datakirjat, 2000. ISBN 951-98558-0-7.
- ^ Alberch P.: Problems with the Interpretation of Developmental Sequences. Systematic Zoology, , nro 34 (1): 46-58, s. 51. Viitannut Matti Kankaanniemi tutkielmassaan Lukion evoluutio-opetus puntarissa.
- ^ aiheesta lisää:Konvergenssi
- ^ Sylvia S. Mader: Biology, s. 298. 6. painos. Boston: WCB/McGraw-Hill, 1998. Lainannut Pekka Reinikainen kirjassaan Dinosaurusten arvoitus ja Raamattu sivuilla 151-153.
- ^ Evoluutio s. 57-58
- ^ Evoluutio s. 58-61
- ^ Lisätietoja: http://creation.com/embryonic-recapitulation-questions-and-answers Luettu 1.6.2015.
- ^ Evoluutio s. 61-62
- ^ Siegfried Scherer, Reinhard Junker: Evoluutio – kriittinen analyysi, s. 169-170. Datakirjat, 2000. ISBN 951-98558-0-7.
- ^ Siegfried Scherer, Reinhard Junker: Evoluutio – kriittinen analyysi, s. 169. Datakirjat, 2000. ISBN 951-98558-0-7.
- ^ Bollinger, R.R., Barbas, A.S., Bush, E.L., Lin, S.S. & Parker. W. (2007) Biofilms in the large bowel suggest an apparent function of the human vermiform appendix. J. Theoretical Biology. 249: 826-831.
- ^ Refuting Evolution 2 Appendix 1: Common arguments for evolution that have been rejected -CMI Luettu 10.11.2010.
- ^ Siegfried Scherer, Reinhard Junker: Evoluutio – kriittinen analyysi, s. 69. Datakirjat, 2000. ISBN 951-98558-0-7.
- ^ Refuting Evolution 2 Appendix 1: Common arguments for evolution that have been rejected -CMI Luettu 10.11.2010.
- ^ Evoluutio s. 62
- ^ >Siegfried Scherer, Reinhard Junker: Evoluutio – kriittinen analyysi, s. 201. Datakirjat, 2000. 951-98558-0-7.
- ^ Evoluutio s. 68
- ^ Mikko Tuuliranta: [http://www.espoohsrk.fi/assets/txt/evoluutio2009.pdf Synteettisen evoluutioteorian suppea analyysi] (s. 58) 2007, osittain päivitetty 10/2009. Viitattu 4.12.2010.
- ^ Evoluutio s. 71
- ^ Tässä tarkastelussa ei edes oteta huomioon edellä mainittujen havaintoaineistojen tuottamia vaikeuksia, vaan kysymystä tarkastellaan "puhtaalta pöydältä" periaatteellisella tasolla.
- ^ http://luominen.fi/evoluution-kumoaminen-2-luku-6-vaitos-yhtenevaiset-rakenteet-viittaavat-yhteiseen-syntyperaan, Luettu 1.6.2015.
- ^ Evoluutio s. 73
- > 65,0 65,1 http://luominen.fi/evoluution-kumoaminen-2-luku-5-vaitos-jotkin-mutaatioista-ovat-hyodyllisia Luettu 1.6.2015.
- ^ Informaation menettäminen olikin odotettavissa informaatioteorian pohjalta.
- ^ Evoluutio s. 73-74
Mayr voi hyvinkin olla oikeassa siltä osin, ettei evoluutioteorialla ole mitään varteenotettavaa kilpailevaa "luonnollista" eli tarkoituksettomien luonnonilmiöiden ja -prosessien varaan rakennettua selitystä. Olennaisin kysymys ei kuitenkaan perimmältään ole tämä vaan se, voiko evoluutioteoriaa ylipäätään perustellusti pitää elollisen luonnon vakavasti otettavana alkuperäselityksenä. Ellei voi, koko "luonnollisiin selityksiin" rajoittuva naturalistinen paradigma on menettänyt älyllisen uskottavuutensa ja se olisi korvattava toimivammalla selittämistavalla, käytännössä ilmeisesti älyllisellä suunnittelulla. - ^ Evoluutio s. 75
- ^ Morfologia tarkoittaa biologisten organismien anatomian tutkimista.
- ^ Evoluutio s. 76
- ^ Evoluutio s. 86
- ^ Pekka Reinikainen: Unohdettu Genesis. Luominen vai evoluutio, s. 145. Kustannus Oy Uusi Tie, 1991. 2. täydennetty painos 1994. ISBN 951-619-239-4.
- ^ Evoluutio s. 78
- ^ Blinkman, R.T. ”Dissociation of Water Vapor and Evolution of Oxygen in the Terrestrial Atmosphere”, Journal of Geophysical Research, vol. 74, No. 23, 20 October 1969, pp. 5355–5368; viitannut Reinikainen, Pekka, Unohdettu Genesis, s.254.
- ^ Evoluutio s. 79
Mayrin näkemystä voi verrata Michael Behen tiivistelmään abiogeneesitutkimuksen nykytilasta:
"Elämän alkuperätutkimuksen vähittäisen halvaantumisen historia on mitä kiintoisin, mutta tilanpuute estää toistamasta sitä tässä. Riittäköön siis maininta, että nykyisellään elämän alkuperätutkimusten kenttä on rämettynyt keskenään ristiriitaisten mallien sekametelisopaksi; kukin malli on epäuskottava, perustavasti puutteellinen ja kilpailevien mallien kanssa yhteensopimaton. Yksityiskeskusteluissa jopa useimmat evoluutiobiologitkin myöntävät, ettei tieteellä ole tarjota elämän alkamiselle mitään selitystä."
("The story of the slow paralysis of research on life's origin is quite interesting, but space precludes its retelling here. Suffice it to say that at present the field of originoflife studies has dissolved into a cacophony of conflicting models, each unconvincing, seriously incomplete, and incompatible with competing models. In private even most evolutionary biologists will admit that science has no explanation for the beginning of life.")
Molecular Machines: Experimental Support for the Design Inference -artikkeli, To Explain Life -luku - ^ Evoluutio s. 80
Tämä perustelu on puhtaasti naturalistista kehäpäättelyä: koska Mayrin naturalististen perususkomusten mukaan elämä on joka tapauksessa joko syntynyt tarkoituksettomasti sattumalta tai ei mitenkään, sen syntymisen tosiasia on hänen mielestään todiste siitä, että se on syntynyt tarkoituksettomasti sattumalta (koska se siis ei ole jäänyt kokonaan syntymättä), vieläpä suhteellisen helposti (koska siihen ei ole tarvittu kovin kauaa aikaa); niinpä se on periaatteessa toistettavissa laboratorioissakin. Koska tämä päättely on niin välittömästi "kaiken oikean tieteen perustana olevien" naturalististen perususkomusten varaista, laboratoriotyöskentelyn tuloksettomuus ei anna Mayrille mitään aihetta kyseenalaistaa sitä: kokeellinen tutkimustyö voi kyllä kumota yksittäisiä hypoteeseja muttei "varmaa tieteellistä tietoa". Sellainen ajatus, että ihmiselle (luonnonprosesseista puhumattakaan) voi olla mahdotonta toistaa Jumalan tekoja, ei tule miehen mieleenkään (tai ei ainakaan pääse hänen tekstiinsä asti). - ^ Evoluutio s. 87
- > 78,0 78,1 http://creation.com/mitochondria-created-to-energize-us, viitattu 12.5.2013
- ^ http://en.wikipedia.org/w/index.php?title=Mitochondrion&oldid=546557989
- ^ Evoluutio s. 104
- ^ Evoluutio s. 110-111
- ^ Evoluutio s. 111-112
- ^ Evoluutio s. 113-117
- ^ Siegfried Scherer, Reinhard Junker: Evoluutio – kriittinen analyysi, s. 215. Datakirjat, 2000. ISBN 951-98558-0-7.
- ^ Evoluutio s. 118
Siinä määrin kuin väite uusien yllättävienkin löytöjen ja darvinistisen kehyskertomuksen ongelmattomasta yhteensopivuudesta ylipäänsä pätee, se asettaa evoluutioteorian selitysvoiman sitä kyseenalaisemmaksi: teoria tai hypoteesi, joka ennustaa ja selittää mitä hyvänsä "melko hyvin" tai "yhtä hyvin", ei ennusta eikä selitä mitään todella hyvin. Se on kuin muovikassi, joka muuttaa tarvittaessa muotoaan kulloisenkin sisältönsä mukaan mutta ei suojaa mitään kohdetta kolhuilta (vastakohtana esim. tietokonelaukku, joka sopii tietynkokoisen ja -muotoisen herkän laitteen kuljetuksiin erinomaisesti muttei ole kätevä yleiskäyttöisenä kauppakassina).
Varsieväkalaesimerkki osoittaa, että eliöpopulaatioiden ominaisuuksien muuttuminen ajan mittaan, jonka Mayr siis antoi evoluution määritelmäksi, ei ole ainakaan mikään yleinen luonnonlaki, vaan populaatio voi "darvinistisen kehyskertomuksen puitteissa" pysyä ominaisuuksiltaan olennaisesti samanlaisena ainakin "60 miljoonaa vuotta". Tähän verratenhan äskettäisluomisnäkemyskin on itse asiassa varsin varovainen, kun väittää kyseisten populaatioiden pysyneen ominaisuuksiltaan samankaltaisina vain n. 6 000 vuotta.
{kind=link}