Ero sivun ”Eliömaailma rappeutuu (kirja)” versioiden välillä
p →Geneettisen entropian validointi todellisissa populaatioissa: vaihdettu pisteen paikka |
Oikolukua; DNA:n kirjoitusasu on johdonmukaista pitää kautta linjan versaalina |
||
| Rivi 1: | Rivi 1: | ||
'''Eliömaailma rappeutuu – geneettinen entropia ja perimän salaisuus''' on eläkkeellä olevan Cornellin yliopiston genetiikan professorin '''John Sanford'''in kirjoittama kirja, jossa hän käsittelee evoluutioteorian perusmekanismeja, mutaatiota ja luonnonvalintaa ja niiden kykyjä selittää luonnon nykyinen monimuotoisuus. Kirjaa vastaan esitettyä kritiikkiä käsitellään artikkelissa [[Eliömaailma rappeutuu -kirjan kritiikki]]. | '''Eliömaailma rappeutuu – geneettinen entropia ja perimän salaisuus''' on eläkkeellä olevan Cornellin yliopiston genetiikan professorin '''John Sanford'''in kirjoittama kirja, jossa hän käsittelee evoluutioteorian perusmekanismeja, mutaatiota ja luonnonvalintaa, ja niiden kykyjä selittää luonnon nykyinen monimuotoisuus. Kirjaa vastaan esitettyä kritiikkiä käsitellään artikkelissa [[Eliömaailma rappeutuu -kirjan kritiikki]]. | ||
== Luku 1: Genomi on elämän kirja, mistä se on tullut? == | == Luku 1: Genomi on elämän kirja, mistä se on tullut? == | ||
Kunkin eliön geeniperimä sisältää kaiken eliön elämäänsä tarvitseman informaation DNA-koodissa. Dna koostuu pienistä | Kunkin eliön geeniperimä sisältää kaiken eliön elämäänsä tarvitseman informaation DNA-koodissa. Dna koostuu pienistä nukleotideiksi kutsutuista molekyyleistä, joita kuvataan kirjaimilla A, C, T, ja G. Ihmisellä tämän koodi käsittää kaksi kolmen miljardin mittaista sarjaa. Tämän suoraviivaisen informaation lisäksi geeniperimässä on monia silmukoita ja haaroja vähän niin kuin tietokoneohjelmassa. Jotkin genomin geenit säätelevät geenejä, jotka edelleen säätelevät muita geenejä. Dna onkin siis moniulotteinen ja itsesäätelevä monimutkainen kokonaisuus, jonka rinnalle ei riitä mikään ihmisen tuottama teknologia vertailukohdaksi. Kaiken lisäksi tämä informaatio on pakattu äärimmäisen pieneen tilaan solun tumaan. | ||
Olennaisin kysymys DNA:oon liittyen on, mistä sen informaatio on tullut ja miten se pysyy olemassa. Evoluutioteorian mukaan selitys on mutaatio ja luonnonvalinta. Mutaation ja luonnonvalinnan toimintaa voidaan kuvata pienen leikkivaunun valmistusprosessilla, jossa ensimmäisen leikkivaunun valmistusohje kopioidaan sokeasti kirjain kerrallaan seuraavan vaunun valmistamiseksi. Vähitellen vaunujen valmistusohjeisiin tulee pieniä kopioijan tekemiä virheitä ja koska kukin valmistusohje kopioidaan aina edeltävästä seuraavan vaunun rakentamiseksi, virheet kerääntyvät. Valmistusohjeet hajoavat ja samalla hajoavat niiden perusteella tehtävät vaunut. | Olennaisin kysymys DNA:oon liittyen on, mistä sen informaatio on tullut ja miten se pysyy olemassa. Evoluutioteorian mukaan selitys on mutaatio ja luonnonvalinta. Mutaation ja luonnonvalinnan toimintaa voidaan kuvata pienen leikkivaunun valmistusprosessilla, jossa ensimmäisen leikkivaunun valmistusohje kopioidaan sokeasti kirjain kerrallaan seuraavan vaunun valmistamiseksi. Vähitellen vaunujen valmistusohjeisiin tulee pieniä kopioijan tekemiä virheitä ja koska kukin valmistusohje kopioidaan aina edeltävästä seuraavan vaunun rakentamiseksi, virheet kerääntyvät. Valmistusohjeet hajoavat ja samalla hajoavat niiden perusteella tehtävät vaunut. | ||
| Rivi 13: | Rivi 13: | ||
== Luku 2: Ovatko sattumanvaraiset mutaatiot hyviä? == | == Luku 2: Ovatko sattumanvaraiset mutaatiot hyviä? == | ||
Mutaatiot saavat aikaan suuria tragedioita | Mutaatiot saavat aikaan suuria tragedioita; esimerkiksi syövät ovat mutaatioiden seurausta. Valtioiden terveyspolitiikat keskittyvätkin mutaatioiden minimoimiseen. Evoluutioteorian mukaan mutaatiot ovat kuitenkin hyviä, sillä ne luovat geneettistä muuntelua. | ||
Muuntelua on kuitenkin kahdenlaista: suunniteltua ja satunnaista. Suunniteltu muuntelu on esimerkiksi autojen lisävarusteet: niiden avulla voidaan saada aikaan muuntelua autossa. Monien eliöiden perimässä on nykyään ihmisen suunnittelemaa geneettistä vaihtelua. Satunnainen muuntelu on taas autossa ruostetta, hajonneita osia ja kiveniskemiä. | Muuntelua on kuitenkin kahdenlaista: suunniteltua ja satunnaista. Suunniteltu muuntelu on esimerkiksi autojen lisävarusteet: niiden avulla voidaan saada aikaan muuntelua autossa. Monien eliöiden perimässä on nykyään ihmisen suunnittelemaa geneettistä vaihtelua. Satunnainen muuntelu on taas autossa ruostetta, hajonneita osia ja kiveniskemiä. | ||
| Rivi 31: | Rivi 31: | ||
# Tämä on myös havaittu tutkimuksissa, joissa on tehty jokin tietty geenimuutos ja tarkkailtu sen vaikutuksia <ref>Sanford, s. 25</ref>. | # Tämä on myös havaittu tutkimuksissa, joissa on tehty jokin tietty geenimuutos ja tarkkailtu sen vaikutuksia <ref>Sanford, s. 25</ref>. | ||
Täysin neutraaleja ne eivät kuitenkaan ole, sillä ENCODE-projektissa<ref name="encode2012">{{Lehtiviite | Tekijä = The ENCODE Project Consortium | Otsikko = An integrated encyclopedia of DNA elements in the human genome | Julkaisu = Nature | Ajankohta = 2012 | Numero = 489 | Sivut = 57-74 | Tunniste = | Viitattu = }}</ref> havaittiin, että perimä on 80%:sti toiminnallista. On olemassa teoreettisia perusteita sen puolesta, että loputkin perimästä olisi toiminnallista. Esimerkiksi olemassaolollaan yksittäinen dna-koodin kirjain vie tilaa ja kuluttaa energiaa solunjakautumisessa sen lisäksi, että se vaikuttaa | Täysin neutraaleja ne eivät kuitenkaan ole, sillä ENCODE-projektissa<ref name="encode2012">{{Lehtiviite | Tekijä = The ENCODE Project Consortium | Otsikko = An integrated encyclopedia of DNA elements in the human genome | Julkaisu = Nature | Ajankohta = 2012 | Numero = 489 | Sivut = 57-74 | Tunniste = | Viitattu = }}</ref> havaittiin, että perimä on 80%:sti toiminnallista. On olemassa teoreettisia perusteita sen puolesta, että loputkin perimästä olisi toiminnallista. Esimerkiksi olemassaolollaan yksittäinen dna-koodin kirjain vie tilaa ja kuluttaa energiaa solunjakautumisessa sen lisäksi, että se vaikuttaa DNA:n toimintaan ja rakenteeseen. Eyre-Walker ja Keightley<ref>{{Lehtiviite | Tekijä = Eyre-Walker, A. ja Keightley, P.D. | Otsikko = The distribution of fitness effects of new mutations | Julkaisu = Nat Rev Genet | Ajankohta = 2007 | Numero = 8 | Sivut = 610-618 | Tunniste = | Viitattu = }} </ref> ovat myös todenneet, että kaikilla mutaatioilla täytyy olla vaikutusta. | ||
Mutaatiot vaihtelevat siis tappavista hyödyllisiin. Naiivi näkemys on, että mutaatioiden vaikutus on jakautunut normaalijakautuneesti niin, että neutraalin raja on jakauman keskellä. Tällaisen jakauman oletti esimerkiksi '''R. A. Fisher''', luodessaan perustavaa luonnonvalinnan teoriaansa. Se on kuitenkin ilmiselvästi väärä, sillä hyödylliset muutokset ovat niin harvinaisia, että ne jätetään tällaisista kaavioista monesti pois. Yksi esimerkki tällaisesta on Kimuran<ref name="kimura79">{{Lehtiviite | Tekijä = Kimura M. | Otsikko = Model of effective neutral mutations in which selective constraint is incorporated | Julkaisu = PNAS | Ajankohta = 1979 | Numero = 76 | Sivut = 3440-3444 | Tunniste = | Viitattu = }}</ref> kaaviosta muokattu<ref>https://luominen.fi/css_images/kimuras_distribution_question.png</ref>, jossa negatiivisten mutaatioiden jakauma lähenee asymptoottisesti kasvaen vaikutuksen nollalinjaa kohti, ja hyödyllisiä ei ole ollenkaan. | |||
<!-- <ref name="kimura79" /> --> | <!-- <ref name="kimura79" /> --> | ||
| Rivi 54: | Rivi 54: | ||
== Luku 3: Miten monta mutaatiota on liian monta? == | == Luku 3: Miten monta mutaatiota on liian monta? == | ||
Jos vahingollisia mutaatioita kertyy yksi ihmistä kohden sukupolvessa, pitkän aikavälin rappeutuminen on varmaa.<ref>{{Lehtiviite | Tekijä = Muller, H.J. | Otsikko = Our load of mutations | Julkaisu = Amer. J. Human Genetics | Ajankohta = 1950 | Numero = 2 | Sivut = 111-176 }} </ref> Näin siksi, että valinnan on poistettava mutaatiot sitä mukaa kun niitä tulee, eikä | Jos vahingollisia mutaatioita kertyy yksi ihmistä kohden sukupolvessa, pitkän aikavälin rappeutuminen on varmaa.<ref>{{Lehtiviite | Tekijä = Muller, H.J. | Otsikko = Our load of mutations | Julkaisu = Amer. J. Human Genetics | Ajankohta = 1950 | Numero = 2 | Sivut = 111-176 }} </ref> Näin siksi, että valinnan on poistettava mutaatiot sitä mukaa kun niitä tulee, eikä valinnalliseen karsimiseen ole käytettävissä kuin osa populaatiosta. Genetiikan kirjallisuudessa kerrottujen arvioiden mukaan kukin ihminen saa syntyessään keskimäärin 75–175 uutta mutaatiota perimäänsä<ref name="lynch2010">{{Lehtiviite | Tekijä = Lynch, M. | Otsikko = Rate, molecular spectrum, and consequences of human mutation | Julkaisu = PNAS | Ajankohta = 2010 | Numero = 107 (3) | Sivut = 961-968 | Tunniste = | Viitattu = }} </ref>{{,}}<ref>{{Lehtiviite | Tekijä = Nachman, M.W. ja Crowell, S.L. | Otsikko = Estimate of the mutation rate per nucleotide in humans | Julkaisu = Genetics | Ajankohta = 2000 | Numero =156 | Sivut = 297-304 | Tunniste = | Viitattu = }} </ref>{{,}}<ref name="kondrashov2002">{{Lehtiviite | Tekijä = Kondrashov, A.S. | Otsikko = Direct Estimate of human per nucleotide mutation rates at 20 loci causing Mendelian diseases | Julkaisu = Human Mutation | Ajankohta = 2002 | Numero = 21 | Sivut = 12-27 | Tunniste = | Viitattu = }} </ref>{{,}}<ref>{{Lehtiviite | Tekijä = Campbell, C.D. ja Eichler. E.E. | Otsikko = Properties and rates of germline mutations in humans | Julkaisu = Trends in Genetics | Ajankohta = 2013 | Numero = 29 | Sivut = 575-584 | Tunniste = | Viitattu = }}</ref>{{,}}<ref>{{Lehtiviite | Tekijä = Xue, Y. ym. | Otsikko = Human Y chromosome base-substitution mutation rate measured by direct sequencing in a deep-rooting pedigree | Julkaisu = Current Biology | Ajankohta = 2009 | Numero = 19 | Sivut = 1453-1457 | Tunniste = | Viitattu = }}</ref> , mutta tähän on laskettu vain genomiin tulevat pistemutaatiot.<ref>Sanford, s. 36</ref> | ||
<!-- <ref name="kondrashov2002" /> | <!-- <ref name="kondrashov2002" /> | ||
<ref name="lynch2010" /> --> | <ref name="lynch2010" /> --> | ||
| Rivi 62: | Rivi 62: | ||
Tämän lisäksi mutaatioita tulee mikrosateliitti-dna-osiin, joissa ne voivat aiheuttaa vakavia geneettisiä sairauksia.<ref>{{Lehtiviite | Tekijä = Sutherland, G.R. ja R.I. Richards | Otsikko = Simple tandem repeats and human disease | Julkaisu = PNAS | Ajankohta = 1995 | Numero = 92 | Sivut = 3636-3641 | Tunniste = | Viitattu = }} </ref> Näitä mutaatioita tapahtuu keskimäärin yksi yhtä tyypillisissä mutaatiovauhtia laskevissa tutkimuksissa huomioon otettua pistemutaatiota kohden.<ref>{{Lehtiviite | Tekijä = Ellegren, H. | Otsikko = Microsatellite mutations in the germline: implications for evolutionary inference | Julkaisu = TIG | Ajankohta = 2000 | Numero = 16 | Sivut = 551-558 | Tunniste = | Viitattu = }}</ref> | Tämän lisäksi mutaatioita tulee mikrosateliitti-dna-osiin, joissa ne voivat aiheuttaa vakavia geneettisiä sairauksia.<ref>{{Lehtiviite | Tekijä = Sutherland, G.R. ja R.I. Richards | Otsikko = Simple tandem repeats and human disease | Julkaisu = PNAS | Ajankohta = 1995 | Numero = 92 | Sivut = 3636-3641 | Tunniste = | Viitattu = }} </ref> Näitä mutaatioita tapahtuu keskimäärin yksi yhtä tyypillisissä mutaatiovauhtia laskevissa tutkimuksissa huomioon otettua pistemutaatiota kohden.<ref>{{Lehtiviite | Tekijä = Ellegren, H. | Otsikko = Microsatellite mutations in the germline: implications for evolutionary inference | Julkaisu = TIG | Ajankohta = 2000 | Numero = 16 | Sivut = 551-558 | Tunniste = | Viitattu = }}</ref> | ||
Aiempien lisäksi tapahtuu myös suurempia kromosomistomutaatioita, kuten deleetioita ja insertioita. Kondrashovin<ref name="kondrashov2002" /> mukaan näitä tapahtuu neljä kutakin sataa pistemutaatiota kohden. Ne voivat kuitenkin vaikuttaa yhdestä miljoonaan | Aiempien lisäksi tapahtuu myös suurempia kromosomistomutaatioita, kuten deleetioita ja insertioita. Kondrashovin<ref name="kondrashov2002" /> mukaan näitä tapahtuu neljä kutakin sataa pistemutaatiota kohden. Ne voivat kuitenkin vaikuttaa yhdestä miljoonaan nukleotidiin, ja ne voivat poistaa nukleotideja kokonaan. Näin ollen muuttuvien nukleotidien määrä sukupolvessa ihmistä kohden on selvästi suurempi, kuin 204 – ja vieläkin on olemassa inversio- ja translokaatiomutaatioita, sekä konversioita, jotka voivat edelleen kaksinkertaistaa esitetyt luvut. Todelliseksi nukleotidimuutosten määräksi voidaankin perustellusti olettaa yli 1000 ihmistä kohden.<ref>Sanford, s. 42-43</ref> | ||
Oletetaan kuitenkin käytännön vuoksi ja evoluutioteorian eduksi, että mutaatioita tapahtuu 100 ihmistä kohden sukupolvessa. Tällainen muutoksen määrä on erittäin pieni osa koko perimää, mutta kun lasketaan nykyään maapallolla elävät noin seitsemän miljardia ihmistä, ja jokaiselle näistä tulleet 100 uutta muutosta perimässä, on muutosten yhteenlaskettu määrä 700 miljardia. Kolmen miljardin kirjaimen perimässä tämä tarkoittaa, että jokainen mahdollinen nukleotidipaikka on keskimäärin mutatoitunut yli 200 kertaa. Kaikki mahdolliset pistemutaatiot ovat tapahtuneet jo nykyään elävien ihmisten parissa. Näiden mutaatioiden vaikutuksia ei kuitenkaan nähdä vielä, mutta ne tulevat tulevien sukupolvien aikana ilmi. | Oletetaan kuitenkin käytännön vuoksi ja evoluutioteorian eduksi, että mutaatioita tapahtuu 100 ihmistä kohden sukupolvessa. Tällainen muutoksen määrä on erittäin pieni osa koko perimää, mutta kun lasketaan nykyään maapallolla elävät noin seitsemän miljardia ihmistä, ja jokaiselle näistä tulleet 100 uutta muutosta perimässä, on muutosten yhteenlaskettu määrä 700 miljardia. Kolmen miljardin kirjaimen perimässä tämä tarkoittaa, että jokainen mahdollinen nukleotidipaikka on keskimäärin mutatoitunut yli 200 kertaa. Kaikki mahdolliset pistemutaatiot ovat tapahtuneet jo nykyään elävien ihmisten parissa. Näiden mutaatioiden vaikutuksia ei kuitenkaan nähdä vielä, mutta ne tulevat tulevien sukupolvien aikana ilmi. | ||
| Rivi 74: | Rivi 74: | ||
Monien ihmisten mielestä luonnonvalinta on kuitenkin taikasauva. Sen ajatellaan voivan tehdä mitä tahansa. Sanford muistelee myös itsellään olleen naivin yliarvioiva käsitys luonnonvalinnan toiminnasta genomin tasolla, jota on tutkinut vain osa populaatiogeneetikoista. He ovat huomattavan älykkäitä, mutta selvästi evoluutioteoriaan sitoutuneita. Heidän matemaattiset mallinsa ovat monimutkaisia, eivätkä biologit yleensä ymmärrä niitä. Tällaiset mallit kuitenkin perustuvat tiettyihin olettamuksiin, aksioomiin, joiden pätevyys ratkaisee laskelmien toimivuuden. | Monien ihmisten mielestä luonnonvalinta on kuitenkin taikasauva. Sen ajatellaan voivan tehdä mitä tahansa. Sanford muistelee myös itsellään olleen naivin yliarvioiva käsitys luonnonvalinnan toiminnasta genomin tasolla, jota on tutkinut vain osa populaatiogeneetikoista. He ovat huomattavan älykkäitä, mutta selvästi evoluutioteoriaan sitoutuneita. Heidän matemaattiset mallinsa ovat monimutkaisia, eivätkä biologit yleensä ymmärrä niitä. Tällaiset mallit kuitenkin perustuvat tiettyihin olettamuksiin, aksioomiin, joiden pätevyys ratkaisee laskelmien toimivuuden. | ||
Evoluutioteorian perustavinta ongelmaa Sanford kutsuu prinsessa ja nukleotidi -paradoksiksi tunnetun lastensadun mukaan, jossa prinsessan kuninkaallisuus havaittiin siitä, ettei hän pystynyt nukkumaan, kun kolmentoista patjan alle oli pantu herne. Tämä kuvaa yksilön ilmiasun, eli fenotyypin ja yksilön geeniperimän eli genotyypin välistä eroa. Mutaatiot tapahtuvat genotyypissä molekyylitasolla, ja valinnan pitäisi pystyä valitsemaan näitä miljardeja muutoksia. Ainoa, mihin valinta kohdistuu, on kuitenkin eliön lisääntymistodennäköisyys. Luonnonvalinta ei koskaan näe yksittäisiä | Evoluutioteorian perustavinta ongelmaa Sanford kutsuu prinsessa ja nukleotidi -paradoksiksi tunnetun lastensadun mukaan, jossa prinsessan kuninkaallisuus havaittiin siitä, ettei hän pystynyt nukkumaan, kun kolmentoista patjan alle oli pantu herne. Tämä kuvaa yksilön ilmiasun, eli fenotyypin ja yksilön geeniperimän eli genotyypin välistä eroa. Mutaatiot tapahtuvat genotyypissä molekyylitasolla, ja valinnan pitäisi pystyä valitsemaan näitä miljardeja muutoksia. Ainoa, mihin valinta kohdistuu, on kuitenkin eliön lisääntymistodennäköisyys. Luonnonvalinta ei koskaan näe yksittäisiä DNA-koodin kirjaimia. | ||
Tilanne onkin samankaltainen | Tilanne onkin samankaltainen kuin prinsessan, joka yrittäisi lukea mittavia sokeankirjoituksella kirjoitettuja kirjoja lukuisten patjojen läpi. Kaikkien mutaatioiden vaikutus ei toki ole sokeankirjoituspisteen kokoinen, vaan välillä se on merkittävämpi, kuten keilapallon kokoinen – tämä on kuitenkin äärimmäisen harvinaista, ja luonnonvalinnan toiminta tällaisia niin sanottuja keilapallomutaatioita vastaan on ilmeistä. Valtaisa enemmistö mutaatioista vaikuttaa kuitenkin kuin sokeankirjoituksen kirjain. | ||
Nukleotidien ja yksilön välinen maailma ovat hyvin erilaiset. Osa tästä erilaisuudesta tulee koosta. Jos nukleotidista tehdään herneen kokoinen, tulee ihmisestä karkeasti otettuna 16 000 kilometriä pitkä. Yksittäinen nukleotidi ei myöskään suoraan vaikuta kuin tietyn geenin käännökseen, ja sitä kautta mRNA:n tuotantoon. Se vaikuttaa taas tietyn entsyymin määrään, joka vaikuttaa taas tiettyyn aineenvaihduntaväylään. Tämä voi edelleen vaikuttaa jonkin solun osaan, ja solun toiminta kudoksen toimintaan. Tämä voi edelleen vaikuttaa eliöön ja siihen todennäköisyyteen, miten tämä mutaatio siirtyy seuraavalle sukupolvelle. | Nukleotidien ja yksilön välinen maailma ovat hyvin erilaiset. Osa tästä erilaisuudesta tulee koosta. Jos nukleotidista tehdään herneen kokoinen, tulee ihmisestä karkeasti otettuna 16 000 kilometriä pitkä. Yksittäinen nukleotidi ei myöskään suoraan vaikuta kuin tietyn geenin käännökseen, ja sitä kautta mRNA:n tuotantoon. Se vaikuttaa taas tietyn entsyymin määrään, joka vaikuttaa taas tiettyyn aineenvaihduntaväylään. Tämä voi edelleen vaikuttaa jonkin solun osaan, ja solun toiminta kudoksen toimintaan. Tämä voi edelleen vaikuttaa eliöön ja siihen todennäköisyyteen, miten tämä mutaatio siirtyy seuraavalle sukupolvelle. | ||
| Rivi 82: | Rivi 82: | ||
Jokainen toiminnallisuuden taso heikentää mutaation vaikutusta kertaluokkia. Yksilön tasolla tapahtuvan valinnan ja genomissa tapahtuvien muutosten eroa voidaan kuvata uudenlaisella tavalla kehittää yliopiston biokemian oppikirjoja. Siinä jokaiseen kirjaan luodaan kaikenlaisia kirjoitusvirheitä, jotka vastaavat esimerkiksi pistemutaatioita, deleetioita ja insertioita. Tämän jälkeen parhaimmin menestyneiden opiskelijoiden kirjat kopioidaan seuraaville opiskelijoille, ja edelleen niihin tuotetaan vastaavia virheitä. | Jokainen toiminnallisuuden taso heikentää mutaation vaikutusta kertaluokkia. Yksilön tasolla tapahtuvan valinnan ja genomissa tapahtuvien muutosten eroa voidaan kuvata uudenlaisella tavalla kehittää yliopiston biokemian oppikirjoja. Siinä jokaiseen kirjaan luodaan kaikenlaisia kirjoitusvirheitä, jotka vastaavat esimerkiksi pistemutaatioita, deleetioita ja insertioita. Tämän jälkeen parhaimmin menestyneiden opiskelijoiden kirjat kopioidaan seuraaville opiskelijoille, ja edelleen niihin tuotetaan vastaavia virheitä. | ||
Tämä menetelmä ei kuitenkaan tuota parempia oppikirjoja, sillä virheiden merkitys menestyksen kannalta on hyvin pieni kaiken muun rinnalla, | Tämä menetelmä ei kuitenkaan tuota parempia oppikirjoja, sillä virheiden merkitys menestyksen kannalta on hyvin pieni kaiken muun rinnalla, mikä vaikuttaa opiskelijoiden menestykseen (motivaatio, luokkahuoneet, opettajat, rakkauselämä jne.) Parhaat tulokset eivät synny vähävirheisimmän oppikirjan, vaan monien muiden syiden seurauksena. Nämä muut tekijät ovat kohinaa, joka estää kirjoitusvirheiden tehokkaan valinnan. | ||
Jos tämän järjestelmän annetaan toimia pidempään, oppikirjat vioittuvat entisestään ja ennen pitkää oppimistulokset laskevat. Tekstin virheiden ja opiskelijoiden menestyksen välinen yhteys on suurelta osin olematon, eikä siksi oppikirjojen vioittumista voida estää. Tämä kuvaa hyvin evoluutioteorian toimintaperiaatetta – vielä sillä lisäyksellä, että tällainen mekanismi tuotti biokemian oppikirjan alunperinkin. | Jos tämän järjestelmän annetaan toimia pidempään, oppikirjat vioittuvat entisestään ja ennen pitkää oppimistulokset laskevat. Tekstin virheiden ja opiskelijoiden menestyksen välinen yhteys on suurelta osin olematon, eikä siksi oppikirjojen vioittumista voida estää. Tämä kuvaa hyvin evoluutioteorian toimintaperiaatetta – vielä sillä lisäyksellä, että tällainen mekanismi tuotti biokemian oppikirjan alunperinkin. | ||
| Rivi 116: | Rivi 116: | ||
Joillakin lajeilla genetiikasta riippumattomista syistä lisääntyminen epäonnistuu puolella yksilöistä. Näillä lajeilla tarvitaan neljä jälkeläistä kahta yksilöä kohti ainoastaan populaation koon säilyttämiseksi. Lisäksi monet geneettiset ominaisuudet eivät periydy. Jotkin geenit toimivat hyvin tietynlaisena yhdistelmänä, mutta muuten niistä on hienoista haittaa. Tämä on totta heteroosissa ja epistasiassa. Valinta tällaisia muutoksia vastaan menee hukkaan, koska ominaisuudet eivät periydy. Ennen todellista valintaa täytyy maksaa nämä kaikki muut lisääntymishinnat, ja tämänkin jälkeen on varaa karsia vain sen verran, ettei populaation koko laske. | Joillakin lajeilla genetiikasta riippumattomista syistä lisääntyminen epäonnistuu puolella yksilöistä. Näillä lajeilla tarvitaan neljä jälkeläistä kahta yksilöä kohti ainoastaan populaation koon säilyttämiseksi. Lisäksi monet geneettiset ominaisuudet eivät periydy. Jotkin geenit toimivat hyvin tietynlaisena yhdistelmänä, mutta muuten niistä on hienoista haittaa. Tämä on totta heteroosissa ja epistasiassa. Valinta tällaisia muutoksia vastaan menee hukkaan, koska ominaisuudet eivät periydy. Ennen todellista valintaa täytyy maksaa nämä kaikki muut lisääntymishinnat, ja tämänkin jälkeen on varaa karsia vain sen verran, ettei populaation koko laske. | ||
Yleinen biologinen toiminnallisuus, eli kelpoisuus on hyvin heikosti periytyvä ominaisuus. Kimuran mukaan periytyvyys on 0,004 (jos periytyvyys on 0, ominaisuus ei periydy ollenkaan, jos 1, ominaisuus periytyy täysin). Ympäristötekijät määräävätkin kelpoisuuden lähes täysin, ja näin ollen ylivoimainen enemmistö kaikkien muiden lisääntymisen estäneiden tekijöiden jälkeenkin tapahtuvasta valinnasta menee hukkaan. Ylimääräisestä populaatiosta voidaan valita tehokkaasti vain erittäin pieni osa. Jatkossa kuitenkin oletetaan, että koko ylimääräinen populaatio on täysin käytettävissä tehokkaaseen valintaan. | Yleinen biologinen toiminnallisuus, eli kelpoisuus, on hyvin heikosti periytyvä ominaisuus. Kimuran mukaan periytyvyys on 0,004 (jos periytyvyys on 0, ominaisuus ei periydy ollenkaan, jos 1, ominaisuus periytyy täysin). Ympäristötekijät määräävätkin kelpoisuuden lähes täysin, ja näin ollen ylivoimainen enemmistö kaikkien muiden lisääntymisen estäneiden tekijöiden jälkeenkin tapahtuvasta valinnasta menee hukkaan. Ylimääräisestä populaatiosta voidaan valita tehokkaasti vain erittäin pieni osa. Jatkossa kuitenkin oletetaan, että koko ylimääräinen populaatio on täysin käytettävissä tehokkaaseen valintaan. | ||
Valinnan hinnan jättäminen huomiotta on Sanfordin mukaan teoreetikkojen suurimpia virheitä. He olettavat, että voivat käyttää valintaa kuinka paljon tahansa, mikä muistuttaa esimerkiksi lännenelokuvien | Valinnan hinnan jättäminen huomiotta on Sanfordin mukaan teoreetikkojen suurimpia virheitä. He olettavat, että voivat käyttää valintaa kuinka paljon tahansa, mikä muistuttaa esimerkiksi lännenelokuvien šeriffiä, joka pystyy ampumaan kuudestilaukeavallaan moninkertaisesti useamman kerran. Teoreetikoilla ei kuitenkaan ole lännenelokuvien käsikirjoittajien tapaan taiteilijan vapautta, vaan heidän on kirjanpitäjän tavoin pidettävä kirjaa käytetystä valinnasta ja sen hinnoista, jotta ne voidaan maksaa. | ||
=== 2. Näkymättömät mutaatiot === | === 2. Näkymättömät mutaatiot === | ||
| Rivi 126: | Rivi 126: | ||
=== 3. Lisääntymiskykyisten lisääntymisen järjestelmällinen estäminen === | === 3. Lisääntymiskykyisten lisääntymisen järjestelmällinen estäminen === | ||
Yksikään yhteiskunta maailmassa ei pyri hallitsemaan tarkoin ihmisten lisääntymistä. | Yksikään yhteiskunta maailmassa ei pyri hallitsemaan tarkoin ihmisten lisääntymistä. Natsi-Saksan kokeilun seuraukset olivat katastrofaaliset, eivätkä nykyiset syntyvyyden säännöstelyohjelmatkaan ole olleet tehokkaita mutaatioiden poistamiseksi. Ihmisten lisääntyminen on enimmäkseen satunnaista, ja tähän tekee poikkeuksen vain tapaukset, joissa mutaatio tai useampi aiheuttaa selvän geneettisen vian. | ||
Ihmispopulaatiosta emme siis voi valita yhtäkään pistemutaatiota. Vaikka näin voitaisiin tehdä, eli mutaatiot voitaisiin havaita ja mutantteja estää lisääntymästä, seuraa nopeasti valinnan hinnan tuottama raja, jota enempää ei voida valita. Tästä tulee erityisen merkittävä ongelma silloin, kun valitaan useita mutaatioita vastaan yhtäaikaisesti. Valinta ei olekaan siis alkuunkaan helppoa. | Ihmispopulaatiosta emme siis voi valita yhtäkään pistemutaatiota. Vaikka näin voitaisiin tehdä, eli mutaatiot voitaisiin havaita ja mutantteja estää lisääntymästä, seuraa nopeasti valinnan hinnan tuottama raja, jota enempää ei voida valita. Tästä tulee erityisen merkittävä ongelma silloin, kun valitaan useita mutaatioita vastaan yhtäaikaisesti. Valinta ei olekaan siis alkuunkaan helppoa. | ||
| Rivi 132: | Rivi 132: | ||
Luonnonvalintaa koskee samat ongelmat kuin keinotekoistakin valintaa. Se ei näe näkymättömiä muutoksia, eikä pysty valitsemaan useita mutaatioita vastaan yhtä aikaa valinnan hinnan takia. Keinotekoinen valinta on itse asiassa tehokkaampaa, koska siinä epäkelvot eliöt hävitetään, kun taas luonnonvalinnassa ainoastaan epäkelvon lisääntymistodennäköisyys laskee. | Luonnonvalintaa koskee samat ongelmat kuin keinotekoistakin valintaa. Se ei näe näkymättömiä muutoksia, eikä pysty valitsemaan useita mutaatioita vastaan yhtä aikaa valinnan hinnan takia. Keinotekoinen valinta on itse asiassa tehokkaampaa, koska siinä epäkelvot eliöt hävitetään, kun taas luonnonvalinnassa ainoastaan epäkelvon lisääntymistodennäköisyys laskee. | ||
Valinta | Valinta kylläkin toimii. Se on todellinen luonnossa havaittava ilmiö, jolla on kuitenkin yhtä todelliset rajansa. Sanford pystyi menestyksekkäästi valinnan avulla jalostamaan kasveja ja luonnonvalinta on poistanut ihmispopulaatiostakin kaikkein pahimmat mutaatiot. Luonnonvalinnan toimivuuden sanford tiivistää lauseeseen: | ||
{{lainaus|”Valinta voi joskus toimia geenitasolla, mutta epäonnistuu järjestelmällisesti genomin tasolla.”<ref>Sanford, s. 57</ref>}} | {{lainaus|”Valinta voi joskus toimia geenitasolla, mutta epäonnistuu järjestelmällisesti genomin tasolla.”<ref>Sanford, s. 57</ref>}} | ||
| Rivi 141: | Rivi 141: | ||
=== 1. Valinnan hinta === | === 1. Valinnan hinta === | ||
Ihmisen geeniperimä käsittää kahtena kopiona kolme miljardia dna-koodin kirjainta. Jokainen on tuonut tähän perimään sata uutta muutosta, mutta perinyt myös kaikki aiemmat muutokset. Tämä tarkoittaa, että jokainen ihminen on mutantti. Kaikkien mutaatioiden valinta samalla tarkoittaisi välitöntä sukupuuttoa. Jo oman sukupolvemme aikana | Ihmisen geeniperimä käsittää kahtena kopiona kolme miljardia dna-koodin kirjainta. Jokainen on tuonut tähän perimään sata uutta muutosta, mutta perinyt myös kaikki aiemmat muutokset. Tämä tarkoittaa, että jokainen ihminen on mutantti. Kaikkien mutaatioiden valinta samalla tarkoittaisi välitöntä sukupuuttoa. Jo oman sukupolvemme aikana ihmiskuntaan on tullut 700 miljardia uutta mutaatiota (7 miljadia ihmistä x 100 mutaatiota). | ||
Ihmisistä voidaan kuitenkin estää lisääntymästä vain alle kolmasosaa, jolloin tämä julma toimenpide poistaisi ihmispopulaatiosta reilut 233 miljardia mutaatiota. Tämänkin jälkeen jäisi jäljelle yli 467 miljardia mutaatiota. Vaikka oletettaisiin, että kaksi kolmasosaa mutaatioista on täysin neutraaleja, jäljellä on silti yli 155 miljardia mutaatiota. Vaikka karsimisen määrä kaksinkertaistettaisiin, eli kahta kolmasosaa ihmisistä estettäisiin lisääntymästä, jäljellä olisi edelleen yli 77 miljardia mutaatiota. Näin voimakkaaseen valintaan meillä ei kuitenkaan ole varaa. | Ihmisistä voidaan kuitenkin estää lisääntymästä vain alle kolmasosaa, jolloin tämä julma toimenpide poistaisi ihmispopulaatiosta reilut 233 miljardia mutaatiota. Tämänkin jälkeen jäisi jäljelle yli 467 miljardia mutaatiota. Vaikka oletettaisiin, että kaksi kolmasosaa mutaatioista on täysin neutraaleja, jäljellä on silti yli 155 miljardia mutaatiota. Vaikka karsimisen määrä kaksinkertaistettaisiin, eli kahta kolmasosaa ihmisistä estettäisiin lisääntymästä, jäljellä olisi edelleen yli 77 miljardia mutaatiota. Näin voimakkaaseen valintaan meillä ei kuitenkaan ole varaa. | ||
| Rivi 147: | Rivi 147: | ||
=== Huomaamattomat ja merkitykseltään vähäiset mutaatiot === | === Huomaamattomat ja merkitykseltään vähäiset mutaatiot === | ||
Tappavat tai lähes tappavat mutaatiot eivät ole populaation näkökulmasta valinnan ongelma, sillä ne ovat harvinaisia ja ne karsiutuvat itsestään. Erityisesti lyhyellä aikavälillä populaation kannalta ongelmallisimpia ovat merkitykseltään vähäiset mutaatiot<ref name="kimurajaohta71" /><sup>s.53</sup>, jotka pitää karsia tai populaatio rappeutuu nopeasti. Vaikka ne saataisiinkin karsittua, jäljellä | Tappavat tai lähes tappavat mutaatiot eivät ole populaation näkökulmasta valinnan ongelma, sillä ne ovat harvinaisia ja ne karsiutuvat itsestään. Erityisesti lyhyellä aikavälillä populaation kannalta ongelmallisimpia ovat merkitykseltään vähäiset mutaatiot<ref name="kimurajaohta71" /><sup>s.53</sup>, jotka pitää karsia tai populaatio rappeutuu nopeasti. Vaikka ne saataisiinkin karsittua, jäljellä ovat edelleen lähes neutraalit mutaatiot. Näin ollen genomit väistämättä rappeutuvat. | ||
==== lähes neutraalit mutaatiot ==== | ==== lähes neutraalit mutaatiot ==== | ||
| Rivi 181: | Rivi 181: | ||
Koko genomi koostuu suurista yhteenlinkittyneistä ryhmistä<ref name="Tishkoff">{{Lehtiviite | Tekijä = Tishkoff, S.A. ja Verrelli, B.C. | Otsikko = Patterns of human genetic diversity: implications for human evolutionary history and disease. | Julkaisu = Annual Review of Genomics and Human Genetics | Ajankohta = 2003 | Numero = 4 | Sivut = 293-340 | Tunniste = | Viitattu = }}</ref>. Jokainen tällainen blokki sisältää vahingollisia muutoksia, vaikka se sisältäisi jonkun merkittävän hyödyllisenkin mutaation. Ryhmän kokonaisvaikutus kelpoisuuteen on siis vahingollinen, vieläpä, kun kuhunkin tällaiseen ryhmään ehtii kertyä tuhansia vahingollisia mutaatioita ennen ensimmäistäkään hyödyllistä. | Koko genomi koostuu suurista yhteenlinkittyneistä ryhmistä<ref name="Tishkoff">{{Lehtiviite | Tekijä = Tishkoff, S.A. ja Verrelli, B.C. | Otsikko = Patterns of human genetic diversity: implications for human evolutionary history and disease. | Julkaisu = Annual Review of Genomics and Human Genetics | Ajankohta = 2003 | Numero = 4 | Sivut = 293-340 | Tunniste = | Viitattu = }}</ref>. Jokainen tällainen blokki sisältää vahingollisia muutoksia, vaikka se sisältäisi jonkun merkittävän hyödyllisenkin mutaation. Ryhmän kokonaisvaikutus kelpoisuuteen on siis vahingollinen, vieläpä, kun kuhunkin tällaiseen ryhmään ehtii kertyä tuhansia vahingollisia mutaatioita ennen ensimmäistäkään hyödyllistä. | ||
Vuonna 2006 julkaistun tutkimuksen<ref name="loewe2006">{{Lehtiviite | Tekijä = Loewe, L. | Otsikko = Quantifying the genome decay paradox due to Muller’s ratchet in human mitochondrial DNA | Julkaisu = Genetic Research | Ajankohta = 2006 | Numero = 87 | Sivut = 133-159 | Tunniste = | Viitattu = }} </ref> mukaan ihmisen olisi pitänyt kuolla sukupuuttoon evolutiivisella aikajanalla. Tutkimuksessa käsiteltiin kuitenkin vain mitokondrion | Vuonna 2006 julkaistun tutkimuksen<ref name="loewe2006">{{Lehtiviite | Tekijä = Loewe, L. | Otsikko = Quantifying the genome decay paradox due to Muller’s ratchet in human mitochondrial DNA | Julkaisu = Genetic Research | Ajankohta = 2006 | Numero = 87 | Sivut = 133-159 | Tunniste = | Viitattu = }} </ref> mukaan ihmisen olisi pitänyt kuolla sukupuuttoon evolutiivisella aikajanalla. Tutkimuksessa käsiteltiin kuitenkin vain mitokondrion DNA:ta, ja se on vain yksi ihmisen 100 000 – 200 000:sta yhdistyneestä ryhmästä. Näin ollen tutkimus aliarvioi yhteenliittymien ongelman jopa 200 000-kertaisesti. | ||
== Luku 6: Kohina == | == Luku 6: Kohina == | ||
| Rivi 199: | Rivi 199: | ||
Jos tarkastellaan yksittäisen nukleotidimuutoksen periytyvyyttä, havaitaan, että sen kelpoisuutta parantava arvo (signaali) on häviävän pieni samalla, kuin kaiken periytymättömän muuntelun ja kaikkien muiden nukleotidipaikkojen yhteisvaikutus (kohina) on tähtitieteellinen. Tästä syystä suurin osa mutaatioista on valinnan ulottumattomissa. | Jos tarkastellaan yksittäisen nukleotidimuutoksen periytyvyyttä, havaitaan, että sen kelpoisuutta parantava arvo (signaali) on häviävän pieni samalla, kuin kaiken periytymättömän muuntelun ja kaikkien muiden nukleotidipaikkojen yhteisvaikutus (kohina) on tähtitieteellinen. Tästä syystä suurin osa mutaatioista on valinnan ulottumattomissa. | ||
Aiempien lisäksi kohinaa tuottaa todennäköisyysvalinta. Siinä missä jalostajat pisteyttävät jalostettavat eliönsä erilaisten parametrien, kuten elinvoiman ja tuottoisuuden mukaan ja sitten määrittelevät rajan, jota heikommat karsitaan täysin ja jota paremmat saavat lisääntyä. Luonnossa näin ei kuitenkaan tapahdu, sillä muutokset saavat aikaan vain eroja lisääntymistodennäköisyyteen ja sattumalta huonommatkin yksilöt saavat lisääntyä. | Aiempien lisäksi kohinaa tuottaa todennäköisyysvalinta. Siinä, missä jalostajat pisteyttävät jalostettavat eliönsä erilaisten parametrien, kuten elinvoiman ja tuottoisuuden mukaan ja sitten määrittelevät rajan, jota heikommat karsitaan täysin ja jota paremmat saavat lisääntyä. Luonnossa näin ei kuitenkaan tapahdu, sillä muutokset saavat aikaan vain eroja lisääntymistodennäköisyyteen ja sattumalta huonommatkin yksilöt saavat lisääntyä. | ||
Luonnossa valinta onkin satunnaista onnekkaimman valintaa, ja siitä puuttuu lähes täysin fenotyypin mukainen järjestelmällinen valinta. Esimerkiksi sammakon lisääntyessä munista ison osan syövät kalat, sammakonpoikasista linnut, vene liiskaa osan ja monet tippuvat vesiputouksesta. Täysikasvuisista sammakoista osa kaivautuu rantatörmään, joka ruoppausoperaation yhteydessä poistetaan. Lisääntymisen epäonnistumisesta jopa puolet johtuu vastaavista fenotyypistä riippumattomista tekijöistä. | Luonnossa valinta onkin satunnaista onnekkaimman valintaa, ja siitä puuttuu lähes täysin fenotyypin mukainen järjestelmällinen valinta. Esimerkiksi sammakon lisääntyessä munista ison osan syövät kalat, sammakonpoikasista linnut, vene liiskaa osan ja monet tippuvat vesiputouksesta. Täysikasvuisista sammakoista osa kaivautuu rantatörmään, joka ruoppausoperaation yhteydessä poistetaan. Lisääntymisen epäonnistumisesta jopa puolet johtuu vastaavista fenotyypistä riippumattomista tekijöistä. | ||
| Rivi 213: | Rivi 213: | ||
== Luku 7: Keinotekoiset pelastusmekanismit == | == Luku 7: Keinotekoiset pelastusmekanismit == | ||
Koska tutkimuksissa on havaittu, että mutaatioita kertyy perimään paljon, on tämä mutaatioiden kertymisnopeusongelma täytynyt ratkaista joillain tavalla. Yksi selitystapa on ollut roska- | Koska tutkimuksissa on havaittu, että mutaatioita kertyy perimään paljon, on tämä mutaatioiden kertymisnopeusongelma täytynyt ratkaista joillain tavalla. Yksi selitystapa on ollut roska-DNA, mutta koska tuoreemmat tutkimukset ovat paljastaneet, että roska-dna:ta ei olekaan – ainakaan yhtään niin paljoa, kuin on väitetty – on keksitty kaksi mekanismia mutaatioiden kertymisongelman ratkaisemiseksi: mutaatioiden määrään perustuva mekanismi ja synergistisen epistasian mekanismi. | ||
Mutaatioiden määrään perustuvan mekanismin mukaan joillain yksilöillä on sattumalta enemmän mutaatioita kuin toisilla, ja kohdistamalla valinta näihin yksilöihin, päästään eroon suuremmasta määrästä mutaatioita pienemmällä valinnan hinnalla. | Mutaatioiden määrään perustuvan mekanismin mukaan joillain yksilöillä on sattumalta enemmän mutaatioita kuin toisilla, ja kohdistamalla valinta näihin yksilöihin, päästään eroon suuremmasta määrästä mutaatioita pienemmällä valinnan hinnalla. | ||
| Rivi 293: | Rivi 293: | ||
=== 9. Melkein kaikki hyödylliset mutaatiot ovat lähes neutraaleja === | === 9. Melkein kaikki hyödylliset mutaatiot ovat lähes neutraaleja === | ||
Lähes neutraalien mutaatioiden näkymättömyys valinnalle ja siitä seuraavat ongelmat on geneetikkojen parissa laajalti hyväksytty, mutta ei sen sijaan sitä, että hyödyllisillä mutaatioilla tämä lähes neutraaliuden ongelma on vielä pahempi. Toivomallamme geenillä on joitain vaikutukseltaan merkittäviä nukleotideja, esimerkiksi entsyymin aktiivisen keskuksen määräävät nukleotidit. Nämäkin ovat turhia ja valinnan ulottumattomissa ilman suuria määriä itsessään hyvin vähäisesti merkittäviä nukleotideja. Näitä nukleotideja ei voida asettaa | Lähes neutraalien mutaatioiden näkymättömyys valinnalle ja siitä seuraavat ongelmat on geneetikkojen parissa laajalti hyväksytty, mutta ei sen sijaan sitä, että hyödyllisillä mutaatioilla tämä lähes neutraaliuden ongelma on vielä pahempi. Toivomallamme geenillä on joitain vaikutukseltaan merkittäviä nukleotideja, esimerkiksi entsyymin aktiivisen keskuksen määräävät nukleotidit. Nämäkin ovat turhia ja valinnan ulottumattomissa ilman suuria määriä itsessään hyvin vähäisesti merkittäviä nukleotideja. Näitä nukleotideja ei voida asettaa eikä pitää paikallaan valinnan kautta, joten ensimmäisestäkään halutusta nukleotidista ei saada kiinni. | ||
=== 10. Vahingolliset mutaatiot takaisin kuvaan === | === 10. Vahingolliset mutaatiot takaisin kuvaan === | ||
| Rivi 303: | Rivi 303: | ||
Mutaatiot eivät myöskään ole täysin satunnaisia, sillä osa genomista mutatoituu paljon useammin kuin toiset osat. Useimmin mutatoituvissa niin sanotuissa kuumissa pisteissä haluamamme mutaatio tulee nopeasti, mutta sillä välin kun odotetaan hitaasti mutatoituvissa paikoissa tapahtuvia muutoksia, ehtivät näissä kuumissa pisteissä nukleotidit vaihtua jo pois haluamistamme. Lisäksi monilla nukleotidipaikoilla on taipumus mutatoitua johonkin tiettyyn nukleotidiin, kuten T. Tämä auttaa silloin, kun T on haluttu, mutta hidastaa, kun A, C tai G on haluttu. Tämä edelleen hidastaa kehittävää valintaa. | Mutaatiot eivät myöskään ole täysin satunnaisia, sillä osa genomista mutatoituu paljon useammin kuin toiset osat. Useimmin mutatoituvissa niin sanotuissa kuumissa pisteissä haluamamme mutaatio tulee nopeasti, mutta sillä välin kun odotetaan hitaasti mutatoituvissa paikoissa tapahtuvia muutoksia, ehtivät näissä kuumissa pisteissä nukleotidit vaihtua jo pois haluamistamme. Lisäksi monilla nukleotidipaikoilla on taipumus mutatoitua johonkin tiettyyn nukleotidiin, kuten T. Tämä auttaa silloin, kun T on haluttu, mutta hidastaa, kun A, C tai G on haluttu. Tämä edelleen hidastaa kehittävää valintaa. | ||
Vuonna 2002 julkaistun tutkimuksen<ref>{{Lehtiviite | Tekijä = Britten, R.J. | Otsikko = Divergence between samples of chimpanzee and human DNA sequences is 5% counting indels | Julkaisu = PNAS | Ajankohta = 2002 | Numero = 99 | Sivut = 13633-13635 | Tunniste = | Viitattu = }} </ref> mukaan ihminen ja simpanssi eroavat noin 150 miljoonassa nukleotidipaikassa 40 miljoonan mutaation seurauksena. Näin ollen sekä simpanssi | Vuonna 2002 julkaistun tutkimuksen<ref>{{Lehtiviite | Tekijä = Britten, R.J. | Otsikko = Divergence between samples of chimpanzee and human DNA sequences is 5% counting indels | Julkaisu = PNAS | Ajankohta = 2002 | Numero = 99 | Sivut = 13633-13635 | Tunniste = | Viitattu = }} </ref> mukaan ihminen ja simpanssi eroavat noin 150 miljoonassa nukleotidipaikassa 40 miljoonan mutaation seurauksena. Näin ollen sekä simpanssi että ihminen ovat saaneet noin 20 miljoonaa mutaatiota kehityslinjallaan, mutta vain noin 1000 näistä on voitu valinnalla levittää populaatioon – loput ovat kertyneet satunnaisen ajautumisen kautta. Vain noin 1000 hyödyllistä muutosta ja noin 20 miljoonaa vahingollista: tämä ei olisi vain tehnyt meistä kantamuotojamme huonompia, vaan olisi tappanut meidät sukupuuttoon. | ||
=== Johtopäätökset === | === Johtopäätökset === | ||
| Rivi 319: | Rivi 319: | ||
Mutaation ja valinnan kyvyttömyys pysäyttää genomin rappeutumista osoittaa evoluutioteorian täysin virheelliseksi – ei vain epäuskottavaksi tai epätodennäköiseksi. Se on vain perusteeton hypoteesi. Tämä johtuu jo siitä, että valinta kohdistuu yksilöön, eikä genomiin. Se ei siis voi estää molekyylitasolla tapahtuvien muutosten aiheuttamaa äärimmäisen monimutkaisten järjestelmien tuhoutumista. On kuin yrittäisi korjata tietokonetta vasaralla. | Mutaation ja valinnan kyvyttömyys pysäyttää genomin rappeutumista osoittaa evoluutioteorian täysin virheelliseksi – ei vain epäuskottavaksi tai epätodennäköiseksi. Se on vain perusteeton hypoteesi. Tämä johtuu jo siitä, että valinta kohdistuu yksilöön, eikä genomiin. Se ei siis voi estää molekyylitasolla tapahtuvien muutosten aiheuttamaa äärimmäisen monimutkaisten järjestelmien tuhoutumista. On kuin yrittäisi korjata tietokonetta vasaralla. | ||
Kirjan alussa käytettiin analogiaa pienestä leluvaunusta, jonka kokoamisohjeita kopioi lähes sokea kirjoitusvirheitä tekevä kirjuri-robotti ja kuinka lähes sokea tuomari-robotti valvoi laatua. Älyllistä ohjausta ei ole. Kysymys on, tuleeko tällaisella tuotantolinjalla autoista avaruusaluksia. Varsin selvää on, ettei tule, vaan vain huonompia autoja. Tuomari-robotti voi vain hidastaa autojen huononemista. Entä jos annettaisiin äärettömästi aikaa? Se tarkoittaisi vain täyttä varmuutta, että tuotantolinjalta ei lopulta tule mitään toimivaa ulos. | Kirjan alussa käytettiin analogiaa pienestä leluvaunusta, jonka kokoamisohjeita kopioi lähes sokea kirjoitusvirheitä tekevä kirjuri-robotti, ja kuviteltiin, kuinka lähes sokea tuomari-robotti valvoi laatua. Älyllistä ohjausta ei ole. Kysymys on, tuleeko tällaisella tuotantolinjalla autoista avaruusaluksia. Varsin selvää on, ettei tule, vaan vain huonompia autoja. Tuomari-robotti voi vain hidastaa autojen huononemista. Entä jos annettaisiin äärettömästi aikaa? Se tarkoittaisi vain täyttä varmuutta, että tuotantolinjalta ei lopulta tule mitään toimivaa ulos. | ||
Intuitiivisesti on selvää, ettei kirjoitusvirheiden kautta tuotantolinjan autoihin tule rakettimoottoria. Vaikka tuomari-robotti näkisikin tällaisen auton, se valitsisi sen pois, sillä rakettimoottori ei tee autosta parempaa autoa. Tuomari-robotti kun ei näe tulevaisuutta, eikä sillä ole käsitystä avaruusaluksesta, ainoa, mitä se voisi tehdä avaruusaluksen aikaansaamiseksi, on valita parempia autoja – ja tämä on paradoksaalista. | Intuitiivisesti on selvää, ettei kirjoitusvirheiden kautta tuotantolinjan autoihin tule rakettimoottoria. Vaikka tuomari-robotti näkisikin tällaisen auton, se valitsisi sen pois, sillä rakettimoottori ei tee autosta parempaa autoa. Tuomari-robotti kun ei näe tulevaisuutta, eikä sillä ole käsitystä avaruusaluksesta, ainoa, mitä se voisi tehdä avaruusaluksen aikaansaamiseksi, on valita parempia autoja – ja tämä on paradoksaalista. | ||
| Rivi 329: | Rivi 329: | ||
Raamatussa kuvataan ajanjakso, jonka aikana ihmiset elivät yli 900-vuotiaiksi ja sisäsiittoisuus oli vaaratonta. Tämä tuntuu aivan järjettömältä, mutta toisaalta ymmärryksemme on rajallista. Emme esimerkiksi tiedä, miksi nisäkkäiden elinikä on tyypillisesti maksimissaan alle 20 vuotta ja ihmisellä noin 120 vuotta. Simpanssillakin maksimielinikä jää alle puoleen 120 vuodesta. Ikääntymisen tiedetään johtuvan mutaatioista, joten jos alun perin mutaatioita ei ollut, ovat yli 900-vuoden eliniät loogisia. | Raamatussa kuvataan ajanjakso, jonka aikana ihmiset elivät yli 900-vuotiaiksi ja sisäsiittoisuus oli vaaratonta. Tämä tuntuu aivan järjettömältä, mutta toisaalta ymmärryksemme on rajallista. Emme esimerkiksi tiedä, miksi nisäkkäiden elinikä on tyypillisesti maksimissaan alle 20 vuotta ja ihmisellä noin 120 vuotta. Simpanssillakin maksimielinikä jää alle puoleen 120 vuodesta. Ikääntymisen tiedetään johtuvan mutaatioista, joten jos alun perin mutaatioita ei ollut, ovat yli 900-vuoden eliniät loogisia. | ||
Nooasta seuraavien sukupolvien aikana näkyy myös, kuinka eliniät laskevat | Nooasta seuraavien sukupolvien aikana näkyy myös, kuinka eliniät laskevat eksponentiaalisesti.<ref>{{Lehtiviite | Tekijä = Holladay, P.M. ja J.M. Watt | Otsikko = De-generation: an exponential decay curve in old testament genealogies | Julkaisu = Evangelical Theological Society Papers | Ajankohta = 2001 | Numero = 2001 | Selite = 52 nd Natl. Conf., Nashville, TN Nov. 15-17, 2000 | Sivut = | Tunniste = | Viitattu = }} </ref> Joko Raamattuun on kuvattu todellisia elinikiä tai sitten Raamatun kirjoittajilla on ollut huomattava tarve esittää ekspotentiaalinen rappeutumiskäyrä ja edistynyttä matematiikkaa sen luomiseksi. Se ei kuitenkaan ole uskottavaa, sillä käytössä ei ollut 1800-luvun havaintoja genetiikasta ja 1900-luvun havaintoja mutaatioista. Lisäksi Sanfordin kollegoineen tekemät simulaatiot tuottavat hyvin tarkasti samanlaisia käyriä. | ||
{{lainaus|”Mikä on genomin salaisuus? Sen pelkkä olemassaolo on sen salaisuus. Ihmisen ymmärryksen ylittävä informaatio ja monimutkaisuus on ohjelmoitu pölyhiukkastakin pienempään tilaan. Mutaatio ja valinta eivät voi alkuunkaan selittää sitä. On täysin selvää, ettei genomimme ole voinut syntyä itsestään. Ainoa perusteltu vaihtoehto itsestään syntyneelle genomille on suunniteltu genomi. Eikö tämä olekin pelottavan suuri salaisuus – tutkistelumme arvoinen?”|Sanford<ref>Sanford,s. 116</ref>}} | {{lainaus|”Mikä on genomin salaisuus? Sen pelkkä olemassaolo on sen salaisuus. Ihmisen ymmärryksen ylittävä informaatio ja monimutkaisuus on ohjelmoitu pölyhiukkastakin pienempään tilaan. Mutaatio ja valinta eivät voi alkuunkaan selittää sitä. On täysin selvää, ettei genomimme ole voinut syntyä itsestään. Ainoa perusteltu vaihtoehto itsestään syntyneelle genomille on suunniteltu genomi. Eikö tämä olekin pelottavan suuri salaisuus – tutkistelumme arvoinen?”|Sanford<ref>Sanford,s. 116</ref>}} | ||
Versio 27. elokuuta 2020 kello 18.53
Eliömaailma rappeutuu – geneettinen entropia ja perimän salaisuus on eläkkeellä olevan Cornellin yliopiston genetiikan professorin John Sanfordin kirjoittama kirja, jossa hän käsittelee evoluutioteorian perusmekanismeja, mutaatiota ja luonnonvalintaa, ja niiden kykyjä selittää luonnon nykyinen monimuotoisuus. Kirjaa vastaan esitettyä kritiikkiä käsitellään artikkelissa Eliömaailma rappeutuu -kirjan kritiikki.
Luku 1: Genomi on elämän kirja, mistä se on tullut?
Kunkin eliön geeniperimä sisältää kaiken eliön elämäänsä tarvitseman informaation DNA-koodissa. Dna koostuu pienistä nukleotideiksi kutsutuista molekyyleistä, joita kuvataan kirjaimilla A, C, T, ja G. Ihmisellä tämän koodi käsittää kaksi kolmen miljardin mittaista sarjaa. Tämän suoraviivaisen informaation lisäksi geeniperimässä on monia silmukoita ja haaroja vähän niin kuin tietokoneohjelmassa. Jotkin genomin geenit säätelevät geenejä, jotka edelleen säätelevät muita geenejä. Dna onkin siis moniulotteinen ja itsesäätelevä monimutkainen kokonaisuus, jonka rinnalle ei riitä mikään ihmisen tuottama teknologia vertailukohdaksi. Kaiken lisäksi tämä informaatio on pakattu äärimmäisen pieneen tilaan solun tumaan.
Olennaisin kysymys DNA:oon liittyen on, mistä sen informaatio on tullut ja miten se pysyy olemassa. Evoluutioteorian mukaan selitys on mutaatio ja luonnonvalinta. Mutaation ja luonnonvalinnan toimintaa voidaan kuvata pienen leikkivaunun valmistusprosessilla, jossa ensimmäisen leikkivaunun valmistusohje kopioidaan sokeasti kirjain kerrallaan seuraavan vaunun valmistamiseksi. Vähitellen vaunujen valmistusohjeisiin tulee pieniä kopioijan tekemiä virheitä ja koska kukin valmistusohje kopioidaan aina edeltävästä seuraavan vaunun rakentamiseksi, virheet kerääntyvät. Valmistusohjeet hajoavat ja samalla hajoavat niiden perusteella tehtävät vaunut.
Valmistusohjeiden hajoamista estämään tulee kuitenkin luonnonvalintaa kuvaava laadunvalvontatuomari, joka valitsee suhteelliselta toimintakyvyltään parhaat vaunut seuraavalle kopiointikierrokselle. Huonojen vaunujen valmistusohjeita ei siis kopioidakaan. Joskus harvoin valmistusohjeeseen voi tulla myös muutos, joka on hyödyllinen. Välillä kopioija voi kopioida jonkin kappaleen, lauseen tai sivun kahteen kertaan, jolloin ohjeen koko voi kasvaa. Harmittomat ylimääräiset kopiot pääsevät tuomarin siivilästä läpi ja niihin saattaa kirjoitusvirheiden kautta kertyä uusien vaununosien informaatiota.
Näin meillä on mekanismi sille, kuinka pienestä leikkivaunusta voi ajan mittaan kehittyä avaruusalus. Avaruusaluskin on kuitenkin riittämätön kuvaamaan ihmisen monimutkaisuutta, joten analogia ei mene riittävän pitkälle. Olennaista on huomata, ettei tässä prosessissa älykkyyttä ole missään. Laadunvalvontatuomarilla ei ole tietoa tulevasta, sillä se kuvaa vain sitä, että jotkin yksilöt lisääntyvät enemmän kuin toiset.
Luku 2: Ovatko sattumanvaraiset mutaatiot hyviä?
Mutaatiot saavat aikaan suuria tragedioita; esimerkiksi syövät ovat mutaatioiden seurausta. Valtioiden terveyspolitiikat keskittyvätkin mutaatioiden minimoimiseen. Evoluutioteorian mukaan mutaatiot ovat kuitenkin hyviä, sillä ne luovat geneettistä muuntelua.
Muuntelua on kuitenkin kahdenlaista: suunniteltua ja satunnaista. Suunniteltu muuntelu on esimerkiksi autojen lisävarusteet: niiden avulla voidaan saada aikaan muuntelua autossa. Monien eliöiden perimässä on nykyään ihmisen suunnittelemaa geneettistä vaihtelua. Satunnainen muuntelu on taas autossa ruostetta, hajonneita osia ja kiveniskemiä.
Satunnainen muuntelu, eli genomissa mutaatiot, ovat lähes aina vahingollista. Tämä näkyy hyödyllisten mutaatioiden äärimmäisenä harvinaisuutena, vaikka tutkijat ovatkin erittäin tarkkana näiden muutosten havaitsemiseksi. Merkittävimpiin esimerkkeihin kuuluvat antibiooteille vastustuskykyiset bakteeritkaan eivät ole kehittyneet, sillä tämän vastustuskyvyn aikaansaavat kromosomistomutaatiot vioittavat solun toimintoja, ja nämä bakteerit häviävät saman tien kilpailussa antibiooteista talttuville bakteereille, kun antibiootti poistetaan ympäristöstä.
Sanford ottaa esimerkiksi kuvitteellisen lentokoneen kokoamisohjeen osan:
Vaihe 6. Kun edellinen vaihe on valmis, toista vaihe 3, kunnes B-osa on 10,004 mm paksu. Sitten odota vähintään 3h, ennen kuin seuraavaan vaiheeseen voidaan siirtyä.
Tähän ohjeen osaan voidaan tuottaa yksittäisiä lyöntivirheitä niin, että niillä on kolmenlaisia vaikutuksia. Ensimmäisenä muutos voisi olla täysin merkityksetön, mutta tässä tapauksessa sellaisesta ei ole antaa esimerkkiä. Toiseksi lyöntivirheellä voisi olla hyvin vähäinen vaikutus, niissä kohdissa, joita ei ole alleviivattu. Kolmanneksi muutokset voivat olla merkittäviä – mitä ne olisivat alleviivatuissa kohdissa. Esimerkistä ei selviä, millainen muutos voisi kehittää kokoamisohjetta: mahdollisesti B-osan paksuus voisi muuttua muutamalla millimetrin tuhannesosalla. Tällainen muutos ei kuitenkaan luo uutta informaatiota, vaan vain säätää olemassa olevaa järjestelmää. Lisäksi haitallisia muutoksia olisi tällaista hienoisesti parantavaa muutosta kohdin moninkertaisesti enemmän.
Sama toimii eliöden genomeissa. Ihmisellä perimässä on kolme miljardia muutoskohtaa, mutta suurimmassa osassa näistä kohdista muutokset ovat vaikutukseltaan lähes neutraaleja. Tästä vallitsee populaatiogeneetikkojen parissa yksimielisyys kolmesta syystä:
- Missä tahansa tekstissä suurin osa pienistä muutoksista saa aikaan vain hyvin mitättömän vaikutuksen kokonaisuuden kannalta.
- Ihmisen geeniperimä käsittää 3 miljardia kirjainta: yhden muutos on vain yksi kolmasmiljardisosan muutos koko genomista
- Tämä on myös havaittu tutkimuksissa, joissa on tehty jokin tietty geenimuutos ja tarkkailtu sen vaikutuksia 1.
Täysin neutraaleja ne eivät kuitenkaan ole, sillä ENCODE-projektissa2 havaittiin, että perimä on 80%:sti toiminnallista. On olemassa teoreettisia perusteita sen puolesta, että loputkin perimästä olisi toiminnallista. Esimerkiksi olemassaolollaan yksittäinen dna-koodin kirjain vie tilaa ja kuluttaa energiaa solunjakautumisessa sen lisäksi, että se vaikuttaa DNA:n toimintaan ja rakenteeseen. Eyre-Walker ja Keightley3 ovat myös todenneet, että kaikilla mutaatioilla täytyy olla vaikutusta.
Mutaatiot vaihtelevat siis tappavista hyödyllisiin. Naiivi näkemys on, että mutaatioiden vaikutus on jakautunut normaalijakautuneesti niin, että neutraalin raja on jakauman keskellä. Tällaisen jakauman oletti esimerkiksi R. A. Fisher, luodessaan perustavaa luonnonvalinnan teoriaansa. Se on kuitenkin ilmiselvästi väärä, sillä hyödylliset muutokset ovat niin harvinaisia, että ne jätetään tällaisista kaavioista monesti pois. Yksi esimerkki tällaisesta on Kimuran4 kaaviosta muokattu5, jossa negatiivisten mutaatioiden jakauma lähenee asymptoottisesti kasvaen vaikutuksen nollalinjaa kohti, ja hyödyllisiä ei ole ollenkaan.
Kimuran kaaviossa on myös lähes neutraalien mutaatioiden alue, jossa tapahtuvia mutaatiota valinta ei näe. Ne eivät kuitenkaan ole vaikutukseltaan neutraaleja. Kimura johtaa tämän alueen koon populaation koosta, joka on vain minimiraja. Müller6 on esimerkiksi todennut, että genetiikasta riippumattomat tekijät vaikuttavat siihen, kuinka suuri osa mutaatioista päätyy valinnan ulottumattomiin.
Sanford on todennut, että genetiikan alan kirjallisuudessa arviot hyödyllisten mutaatioiden suhteesta vahingollisiin vaihtelevat välillä 1:1000 – 1:1000 000, niin, että parhaimmat arviot ovat lähempänä 1:1000 000:aa.7, 8. Todellisesta suhteesta ei edes voida tehdä tarkkoja laskelmia sen pienuuden takia.9, 10. Niitä kuitenkin on, mutta ne menevät kaikki Kimuran valinnan ulottumattomissa olevalle alueelle, eikä valinta näe niitä. Joillain äärimmäisen harvoilla hyödyllisillä mutaatioilla on niin merkittävä vaikutus, että valinta näkee ne, ne on kaaviossa merkitty nuolilla. Kaaviosta näkyy selvästi, että informaatiota tuhoavat mutaatiot jyräävät selvästi hyödyllisten yli.
Hyödylliset mutaatiot ovat kuitenkin evoluutioteorian ytimessä, joten viime vuosisadalla käytettiin huomattavia resursseja saamaan mutaatioilla aikaan hyödyllisiä muunnoksia. Kyse on siis mutaatiojalostuksesta, jota alettin harjoittaa sen jälkeen, kun havaittiin tietynlaisen säteilyn ja tiettyjen kemikaalien aiheuttavan mutaatioita. Evoluutioteorian perusteella runsas mutaatioiden määrä ja tehokas valinta kiihdyttäisi kehitystä merkittävästi.
Tämän jälkeen erityisesti kasveja altistettiin säteilylle ja kemikaaleille mutaatioiden aikaansaamiseksi ja sen jälkeen näistä pyrittiin seulomaan mahdollisesti syntyneet paremmat muodot. Valtavia määriä viljelykasveja käsiteltiin näin, ja aikaan saatiin lisääntymiskyvyttömiä, kitukasvuisia, sairaita tai muuten epämuodostuneita kasveja. Sato ei parantunut käytännössä lainkaan, vaikka tohtoritason tiedemiehet olivat ohjaamassa evoluutioprosessia.
Näin kävi, koska mutaatiot eivät tuottaneet hyödyllisiä muutoksia. Pari poikkeusta tällä kuitenkin saavutettiin: vähäfytaattinen maissi ja joitain koristekasveja. Vähäfytaattisella maissilla mutaatiot hajottivat fytaattia valmistavat solujärjestelmät, eikä maissi näin tuottanut enää fytaattia. Tästä oli etua, kun maissia käytetään karjan rehuna, mutta kyse on informaation tuhoutumisesta, ei uuden informaation synnystä. Koristekasvit tarkoittavat monia epämuodostuneita, täplikkäitä tai muuten värivirheellisiä, lisääntymiskyvyttömiä kasveja, joiden epämuodostumia pidettiin kauniina.
Miten evoluutioprosessi eli mutaatio ja valinta voisi toimia ilman älykästä ohjausta, jos se ei toimi edes ohjattuna? Epäonnistuneiden mutaatiojalostuskokeiden jälkeen samat tutkijat siirtyivät perinteiseen jalostukseen, jossa käytetään kasvien luontaista muuntelupotentiaalia. Tällä perinteisellä jalostuksella he saivat merkittäviä saavutuksia aikaan. Kuva on järkevä vain, jos perimän muuntelupotentiaali on käytännössä kokonaan suunniteltua, eikä mutaatioiden aikaansaamaa11 – vastaavalla tavalla kuin autoon voi tuottaa suunniteltua vaihtelua lisävarustein ja suunnittelematonta vaihtelua kiveniskemien, ruostumisen ja hajonneiden osien kautta.12
Bergman13 haki Biological Abstracts and Medlinestä tutkimuksia mutaatio-hakusanalla. Näitä löytyi yli 400 000, mutta vain 186 näistä käsitti sanan benefical – hyödyllinen. Näissäkin kaikissa oli kyse informaatiota tuhonneista muutoksista, jotka olivat hyödyllisiä. Joissain tutkimuksissa on havaittu suuria määriä hyödyllisiä mutaatioita, mutta näissä on kyse tasaisissa laboratorio-olosuhteissa hyödyttömien genomin osien tuhoutumisesta. Monien mikrobien perimästä jopa puolet koostuu geeneistä, joita tarvitaan vain tietynlaisessa elinympäristössä, joita ei laboratoriossa tarvita.14
Laboratorio-olosuhteiden vaikutus näkyy Lenskin tutkimusryhmän tuloksista.15, 16 E. Coli -bakteerit sopeutuivat keinotekoiseen ympäristöön ja kasvoivat siinä hieman nopeammin. Tämä etu tuli kuitenkin siitä, että geenit tuhoutuivat. Bataillonin ja Baileyn17 tutkimus myös tukee mainittua mutaatioiden vaikutuksen totuudenmukaista jakaumaa.
Luku 3: Miten monta mutaatiota on liian monta?
Jos vahingollisia mutaatioita kertyy yksi ihmistä kohden sukupolvessa, pitkän aikavälin rappeutuminen on varmaa.18 Näin siksi, että valinnan on poistettava mutaatiot sitä mukaa kun niitä tulee, eikä valinnalliseen karsimiseen ole käytettävissä kuin osa populaatiosta. Genetiikan kirjallisuudessa kerrottujen arvioiden mukaan kukin ihminen saa syntyessään keskimäärin 75–175 uutta mutaatiota perimäänsä19, 20, 21, 22, 23 , mutta tähän on laskettu vain genomiin tulevat pistemutaatiot.24
Pistemutaatioiden lisäksi mutaatioita tulee mitokondrioihin, joihin niitä on arvoitu tulevan 2,5 kappaletta nukleotidiasemaa kohti miljoonassa vuodessa25. Koska mitokondrion perimä on 16 500 nukleotidia pitkä, ja jos oletetaan 25 vuoden sukupolvi, tämä tarkoittaa yhtä mutaatiota mitokondrion perimässä ihmistä kohden.
Tämän lisäksi mutaatioita tulee mikrosateliitti-dna-osiin, joissa ne voivat aiheuttaa vakavia geneettisiä sairauksia.26 Näitä mutaatioita tapahtuu keskimäärin yksi yhtä tyypillisissä mutaatiovauhtia laskevissa tutkimuksissa huomioon otettua pistemutaatiota kohden.27
Aiempien lisäksi tapahtuu myös suurempia kromosomistomutaatioita, kuten deleetioita ja insertioita. Kondrashovin21 mukaan näitä tapahtuu neljä kutakin sataa pistemutaatiota kohden. Ne voivat kuitenkin vaikuttaa yhdestä miljoonaan nukleotidiin, ja ne voivat poistaa nukleotideja kokonaan. Näin ollen muuttuvien nukleotidien määrä sukupolvessa ihmistä kohden on selvästi suurempi, kuin 204 – ja vieläkin on olemassa inversio- ja translokaatiomutaatioita, sekä konversioita, jotka voivat edelleen kaksinkertaistaa esitetyt luvut. Todelliseksi nukleotidimuutosten määräksi voidaankin perustellusti olettaa yli 1000 ihmistä kohden.28
Oletetaan kuitenkin käytännön vuoksi ja evoluutioteorian eduksi, että mutaatioita tapahtuu 100 ihmistä kohden sukupolvessa. Tällainen muutoksen määrä on erittäin pieni osa koko perimää, mutta kun lasketaan nykyään maapallolla elävät noin seitsemän miljardia ihmistä, ja jokaiselle näistä tulleet 100 uutta muutosta perimässä, on muutosten yhteenlaskettu määrä 700 miljardia. Kolmen miljardin kirjaimen perimässä tämä tarkoittaa, että jokainen mahdollinen nukleotidipaikka on keskimäärin mutatoitunut yli 200 kertaa. Kaikki mahdolliset pistemutaatiot ovat tapahtuneet jo nykyään elävien ihmisten parissa. Näiden mutaatioiden vaikutuksia ei kuitenkaan nähdä vielä, mutta ne tulevat tulevien sukupolvien aikana ilmi.
Kun mutaatioita kertyy paljon, se johtaa lopulta mutaatiosulamiseksi29 kutsuttuun ilmiöön, jossa populaation hedelmällisyys laskee, ja samalla valinnan vaikutusmahdollisuudet edelleen vähenevät. Valinta vaatii ylimääräistä populaatiota, josta voi karsia. Lopulta edessä on sukupuutto. Se uhkaa jo monia nykyisin uhanalaisia lajeja, ja samoin ihmistä vähän pidemmällä aikajänteellä.
Luku 4: luonnonvalinta
Vaikka ihmisten eliniät kasvavatkin yhteiskunnassa, mutaatiot rappeuttavat perimää nopeasti. Tästä vallitsee geneetikkojen parissa yksimielisyys, sillä mutaatioita kasautuu nopeasti ja valinta on heikkoa.30, 19
Monien ihmisten mielestä luonnonvalinta on kuitenkin taikasauva. Sen ajatellaan voivan tehdä mitä tahansa. Sanford muistelee myös itsellään olleen naivin yliarvioiva käsitys luonnonvalinnan toiminnasta genomin tasolla, jota on tutkinut vain osa populaatiogeneetikoista. He ovat huomattavan älykkäitä, mutta selvästi evoluutioteoriaan sitoutuneita. Heidän matemaattiset mallinsa ovat monimutkaisia, eivätkä biologit yleensä ymmärrä niitä. Tällaiset mallit kuitenkin perustuvat tiettyihin olettamuksiin, aksioomiin, joiden pätevyys ratkaisee laskelmien toimivuuden.
Evoluutioteorian perustavinta ongelmaa Sanford kutsuu prinsessa ja nukleotidi -paradoksiksi tunnetun lastensadun mukaan, jossa prinsessan kuninkaallisuus havaittiin siitä, ettei hän pystynyt nukkumaan, kun kolmentoista patjan alle oli pantu herne. Tämä kuvaa yksilön ilmiasun, eli fenotyypin ja yksilön geeniperimän eli genotyypin välistä eroa. Mutaatiot tapahtuvat genotyypissä molekyylitasolla, ja valinnan pitäisi pystyä valitsemaan näitä miljardeja muutoksia. Ainoa, mihin valinta kohdistuu, on kuitenkin eliön lisääntymistodennäköisyys. Luonnonvalinta ei koskaan näe yksittäisiä DNA-koodin kirjaimia.
Tilanne onkin samankaltainen kuin prinsessan, joka yrittäisi lukea mittavia sokeankirjoituksella kirjoitettuja kirjoja lukuisten patjojen läpi. Kaikkien mutaatioiden vaikutus ei toki ole sokeankirjoituspisteen kokoinen, vaan välillä se on merkittävämpi, kuten keilapallon kokoinen – tämä on kuitenkin äärimmäisen harvinaista, ja luonnonvalinnan toiminta tällaisia niin sanottuja keilapallomutaatioita vastaan on ilmeistä. Valtaisa enemmistö mutaatioista vaikuttaa kuitenkin kuin sokeankirjoituksen kirjain.
Nukleotidien ja yksilön välinen maailma ovat hyvin erilaiset. Osa tästä erilaisuudesta tulee koosta. Jos nukleotidista tehdään herneen kokoinen, tulee ihmisestä karkeasti otettuna 16 000 kilometriä pitkä. Yksittäinen nukleotidi ei myöskään suoraan vaikuta kuin tietyn geenin käännökseen, ja sitä kautta mRNA:n tuotantoon. Se vaikuttaa taas tietyn entsyymin määrään, joka vaikuttaa taas tiettyyn aineenvaihduntaväylään. Tämä voi edelleen vaikuttaa jonkin solun osaan, ja solun toiminta kudoksen toimintaan. Tämä voi edelleen vaikuttaa eliöön ja siihen todennäköisyyteen, miten tämä mutaatio siirtyy seuraavalle sukupolvelle.
Jokainen toiminnallisuuden taso heikentää mutaation vaikutusta kertaluokkia. Yksilön tasolla tapahtuvan valinnan ja genomissa tapahtuvien muutosten eroa voidaan kuvata uudenlaisella tavalla kehittää yliopiston biokemian oppikirjoja. Siinä jokaiseen kirjaan luodaan kaikenlaisia kirjoitusvirheitä, jotka vastaavat esimerkiksi pistemutaatioita, deleetioita ja insertioita. Tämän jälkeen parhaimmin menestyneiden opiskelijoiden kirjat kopioidaan seuraaville opiskelijoille, ja edelleen niihin tuotetaan vastaavia virheitä.
Tämä menetelmä ei kuitenkaan tuota parempia oppikirjoja, sillä virheiden merkitys menestyksen kannalta on hyvin pieni kaiken muun rinnalla, mikä vaikuttaa opiskelijoiden menestykseen (motivaatio, luokkahuoneet, opettajat, rakkauselämä jne.) Parhaat tulokset eivät synny vähävirheisimmän oppikirjan, vaan monien muiden syiden seurauksena. Nämä muut tekijät ovat kohinaa, joka estää kirjoitusvirheiden tehokkaan valinnan.
Jos tämän järjestelmän annetaan toimia pidempään, oppikirjat vioittuvat entisestään ja ennen pitkää oppimistulokset laskevat. Tekstin virheiden ja opiskelijoiden menestyksen välinen yhteys on suurelta osin olematon, eikä siksi oppikirjojen vioittumista voida estää. Tämä kuvaa hyvin evoluutioteorian toimintaperiaatetta – vielä sillä lisäyksellä, että tällainen mekanismi tuotti biokemian oppikirjan alunperinkin.
Prinsessa ja nukleotidi -paradoksia pahentaa edelleen homeostaasi-ilmiö, joka tarkoittaa sitä, että eliöt sopeutuvat erilaisiin ympäristöihin, esimerkiksi tasalämpöiset eläimet kylmään ilmastoon. Homeostaasi vaatii hyvin monimutkaista tunnistimien ja säätimien järjestelmää jokaisessa solussa, ja sen seurauksena yksittäisten mutaatioiden vaikutus edelleen vähenee. Tähän vertautuu se, että prinsessan patjat olisivat itseoikaisevia – vaikka patjojen alle pantaisiin tennispallo, ne oikaisisivat itsensä niin, ettei sitä pystyisi havaitsemaan.
Kun Mendelin periaatteet jälleen löydettiin 50 vuotta Mendelin jälkeen, havaittiin eliöissä olevan lukuisia toistaan erillisiä geneettisiä yksiköitä, ja että valinta on ongelmissa, jos näiden yksiköiden määrä kasvaa kovin suureksi. Tämä on prinsenssa ja nukleotidi -paradoksi, vaikka sillä nimellä ei tätä luonnollisesti kutsuttukaan.
Niinpä tutkijat keksivät määritellä populaatiot geenipooleiksi, jossa geenit ovat kuin varastossa ja valinta valitsee niistä parhaita yksitellen. Tämä suoranaisesti pelasti koko evoluutioteorian, mutta on täysin virheellinen: populaatiot ovat erittäin kaukana geenien varastosta.31
Jotta populaation määrittely geenipooliksi onnistuisi, on tutkijoiden täytynyt olettaa useita virheellisiä olettamuksia tosiksi:
- Oletettiin, että kukin nukleotidi valitaan ja peritään yksittäin, eli että genomissa ei ole yhteenliittymiä, joissa useampia nukleotideja perittäisiin kokonaisuuksina. Tällaisia yhteenliittymiä kuitenkin on olemassa.
- Nukleotidien välillä ei ole vuorovaikutusta (epistasia). Tätä on havaittu olevan.
- Populaatioiden koot oletettiin äärettömiksi. Ne eivät kuitenkaan ole sitä ilmiselvästi koskaan.
- Valinnalle oletettiin rajattomasti aikaa. Aikaa on kuitenkin rajallisesti.
- Oletettiin, että rajattomasti ominaisuuksia voidaan valita yhtä aikaa. (Sanford osoittaa tämän virheelliseksi myöhemmin kirjassa. Todellisuudessa vain kourallista ominaisuuksia voidaan valita tehokkaasti yhtä aikaa.)
Nämä olettamukset eivät vastaa biologista todellisuutta, jossa todelliset eliöt elävät. Näillä tutkijoilla oli kuitenkin Sanfordin mukaan valtava ”älyllisen auktoriteetin sädekehä”, jonka varjolla nämä olettamukset hyväksyttiin. Olivathan biologitkin evoluutioteoriaan uskovia, joten heistä oli vain hienoa, että saavat olla samaa mieltä populaatiogenetiikan gurujen kanssa, vaikka eivät pitäneetkään kuvausta populaatioista geenipooleina täysin järkevänä. Sanford itsekin muistelee hyväksyneensä tämän selityksen: hänhän tiesi evoluution olevan totta, joten ei ollut niin tärkeää, vaikka hän ei pitänytkään itseään riittävän fiksuna ymmärtämään populaatioita geenipooleina.32
Todellisuudessa populaatiot eivät ole geenipooleja, vaan geenit ovat sitoutuneet yksilöihin. Genomissa on myös isoja yhteen liittyneitä ryhmiä, joissa on 10 000:sta miljoonaan nukleotidia. Nämä ryhmät eivät hajoa koskaan, vaan niitä käsitellään aina yhtenä kokonaisuutena. Prinsessa ja nukleotidi -paradoksi on siis edelleen ratkaisematta. Se osoittaa yksinään evoluutioteorian virheelliseksi, mutta on vasta ongelmien alkusoittoa. Sanford antaa kuitenkin loppukirjansa ajan teoreetikkojen pitää olettamuksensa populaatioista geenipooleina ja nukleotidien tasolla toimivasta valinnasta.
Perustavimmat valintaongelmat
- Valinnan hinta
- Näkymättömien mutaatioiden havaitseminen
- Järjestelmällinen lisääntymiskykyisten karsinta.
1. Valinnan hinta
Valinnalla on hintansa33, 34, 35, 36, 37. Se voi kohdistua vain ylimääräiseen populaatioon. Esimerkiksi ihmisellä populaation koon ylläpitämiseksi kahden vanhemman on tuotettava yli kaksi jälkeläistä, sillä osa ihmisistä ei lisäänny genetiikasta riippumattomista syistä. Vasta tämän ylittävästä populaatiosta voidaan estää yksilöitä lisääntymästä valinnallisista syistä. Haldanen33 mukaan vain 10% ihmispopulaatiosta voidaan valinnallisesti karsia. Suuremmasta karsimisesta seuraisi nopeasti sukupuutto.
Joillakin lajeilla genetiikasta riippumattomista syistä lisääntyminen epäonnistuu puolella yksilöistä. Näillä lajeilla tarvitaan neljä jälkeläistä kahta yksilöä kohti ainoastaan populaation koon säilyttämiseksi. Lisäksi monet geneettiset ominaisuudet eivät periydy. Jotkin geenit toimivat hyvin tietynlaisena yhdistelmänä, mutta muuten niistä on hienoista haittaa. Tämä on totta heteroosissa ja epistasiassa. Valinta tällaisia muutoksia vastaan menee hukkaan, koska ominaisuudet eivät periydy. Ennen todellista valintaa täytyy maksaa nämä kaikki muut lisääntymishinnat, ja tämänkin jälkeen on varaa karsia vain sen verran, ettei populaation koko laske.
Yleinen biologinen toiminnallisuus, eli kelpoisuus, on hyvin heikosti periytyvä ominaisuus. Kimuran mukaan periytyvyys on 0,004 (jos periytyvyys on 0, ominaisuus ei periydy ollenkaan, jos 1, ominaisuus periytyy täysin). Ympäristötekijät määräävätkin kelpoisuuden lähes täysin, ja näin ollen ylivoimainen enemmistö kaikkien muiden lisääntymisen estäneiden tekijöiden jälkeenkin tapahtuvasta valinnasta menee hukkaan. Ylimääräisestä populaatiosta voidaan valita tehokkaasti vain erittäin pieni osa. Jatkossa kuitenkin oletetaan, että koko ylimääräinen populaatio on täysin käytettävissä tehokkaaseen valintaan.
Valinnan hinnan jättäminen huomiotta on Sanfordin mukaan teoreetikkojen suurimpia virheitä. He olettavat, että voivat käyttää valintaa kuinka paljon tahansa, mikä muistuttaa esimerkiksi lännenelokuvien šeriffiä, joka pystyy ampumaan kuudestilaukeavallaan moninkertaisesti useamman kerran. Teoreetikoilla ei kuitenkaan ole lännenelokuvien käsikirjoittajien tapaan taiteilijan vapautta, vaan heidän on kirjanpitäjän tavoin pidettävä kirjaa käytetystä valinnasta ja sen hinnoista, jotta ne voidaan maksaa.
2. Näkymättömät mutaatiot
Useat tutkijat ovat havainneet38, 39, 40, 41, 42, 43, että suuri osa mutaatioista on näkymättömiä. Ne eivät saa aikaan sellaista vaikutusta, että ne voitaisiin tunnistaa. Tällaisia on suurin osa pistemutaatioista. Tämä tekee myös valinnasta mielivaltaista, sillä se on kuin yrittäisi valita kirjakaupassa tietosanakirjasarjan, josta tietää, että kussakin tietosanakirjasarjasetissä on omat 10 000 kirjoitusvirhettä. Millä tavoin voisi valita vähävirheisimmän tietosanakirjasarjan?
3. Lisääntymiskykyisten lisääntymisen järjestelmällinen estäminen
Yksikään yhteiskunta maailmassa ei pyri hallitsemaan tarkoin ihmisten lisääntymistä. Natsi-Saksan kokeilun seuraukset olivat katastrofaaliset, eivätkä nykyiset syntyvyyden säännöstelyohjelmatkaan ole olleet tehokkaita mutaatioiden poistamiseksi. Ihmisten lisääntyminen on enimmäkseen satunnaista, ja tähän tekee poikkeuksen vain tapaukset, joissa mutaatio tai useampi aiheuttaa selvän geneettisen vian.
Ihmispopulaatiosta emme siis voi valita yhtäkään pistemutaatiota. Vaikka näin voitaisiin tehdä, eli mutaatiot voitaisiin havaita ja mutantteja estää lisääntymästä, seuraa nopeasti valinnan hinnan tuottama raja, jota enempää ei voida valita. Tästä tulee erityisen merkittävä ongelma silloin, kun valitaan useita mutaatioita vastaan yhtäaikaisesti. Valinta ei olekaan siis alkuunkaan helppoa.
Luonnonvalintaa koskee samat ongelmat kuin keinotekoistakin valintaa. Se ei näe näkymättömiä muutoksia, eikä pysty valitsemaan useita mutaatioita vastaan yhtä aikaa valinnan hinnan takia. Keinotekoinen valinta on itse asiassa tehokkaampaa, koska siinä epäkelvot eliöt hävitetään, kun taas luonnonvalinnassa ainoastaan epäkelvon lisääntymistodennäköisyys laskee.
Valinta kylläkin toimii. Se on todellinen luonnossa havaittava ilmiö, jolla on kuitenkin yhtä todelliset rajansa. Sanford pystyi menestyksekkäästi valinnan avulla jalostamaan kasveja ja luonnonvalinta on poistanut ihmispopulaatiostakin kaikkein pahimmat mutaatiot. Luonnonvalinnan toimivuuden sanford tiivistää lauseeseen:
”Valinta voi joskus toimia geenitasolla, mutta epäonnistuu järjestelmällisesti genomin tasolla.”44
Luku 5 Genomitason valinnan ongelmat
Geneetikkojen käytäntönä on Sanfordin mukaan ollut yleistää yhden mutaation valintaa koskevat päätelmät koskemaan kaikkia mutaatioita periaatteella, se mikä toimii yhdellä, toimii kaikilla. Tämä on kuitenkin kuin väittäisi, että jos jollakulla on varaa ostaa yksi auto, hänellä on varaa ostaa miten monta autoa tahansa.
1. Valinnan hinta
Ihmisen geeniperimä käsittää kahtena kopiona kolme miljardia dna-koodin kirjainta. Jokainen on tuonut tähän perimään sata uutta muutosta, mutta perinyt myös kaikki aiemmat muutokset. Tämä tarkoittaa, että jokainen ihminen on mutantti. Kaikkien mutaatioiden valinta samalla tarkoittaisi välitöntä sukupuuttoa. Jo oman sukupolvemme aikana ihmiskuntaan on tullut 700 miljardia uutta mutaatiota (7 miljadia ihmistä x 100 mutaatiota).
Ihmisistä voidaan kuitenkin estää lisääntymästä vain alle kolmasosaa, jolloin tämä julma toimenpide poistaisi ihmispopulaatiosta reilut 233 miljardia mutaatiota. Tämänkin jälkeen jäisi jäljelle yli 467 miljardia mutaatiota. Vaikka oletettaisiin, että kaksi kolmasosaa mutaatioista on täysin neutraaleja, jäljellä on silti yli 155 miljardia mutaatiota. Vaikka karsimisen määrä kaksinkertaistettaisiin, eli kahta kolmasosaa ihmisistä estettäisiin lisääntymästä, jäljellä olisi edelleen yli 77 miljardia mutaatiota. Näin voimakkaaseen valintaan meillä ei kuitenkaan ole varaa.
Huomaamattomat ja merkitykseltään vähäiset mutaatiot
Tappavat tai lähes tappavat mutaatiot eivät ole populaation näkökulmasta valinnan ongelma, sillä ne ovat harvinaisia ja ne karsiutuvat itsestään. Erityisesti lyhyellä aikavälillä populaation kannalta ongelmallisimpia ovat merkitykseltään vähäiset mutaatiot34s.53, jotka pitää karsia tai populaatio rappeutuu nopeasti. Vaikka ne saataisiinkin karsittua, jäljellä ovat edelleen lähes neutraalit mutaatiot. Näin ollen genomit väistämättä rappeutuvat.
lähes neutraalit mutaatiot
Suurin osa mutaatioista on vaikutukseltaan lähes neutraaleja ja niiden vaikutus jää kaikenlaisen biologisen kohinan jalkoihin.38, 35, 34 Näillä mutaatioilla on kuitenkin vaikutusta, sillä ne tuhoavat näissä nukleotidipaikoissa olevaa informaatiota. Ne vastaavat autossa ruostumista, joka tapahtuu atomi kerrallaan. Vaikka hajonneita osia autosta voikin korjata, kaikkialla tapahtuvaa ruostumista ei pysty pysäyttämään. Sama pätee myös ihmisen ikääntymiseen, vaikka elimiä voidaan vaihtaa, vanhenemista aiheuttavia mutaatioita soluissa ei voida pysäyttää. Tämä ongelma on saanut yhden tunnetun populaatiogeneetikon kirjoittamaan tutkimuksen otsikolla "Why have we not died 100 times over?"43.
Homogeenisessa ympäristössä elävässä bakteeripopulaatiossa valinta on tehokasta ja jokainen solu on itsenäisesti valinnan kohde. Niiden genomit ovat pieniä ja geenien väliset vuorovaikutukset suhteellisen vähäisiä. Ne lisääntyvät nopeasti ja mutaatioita tulee vain vähän kuhunkin genomiin. Huomaamattomia mutaatioita on tällaisessa populaatiossa vähän. Ihmispopulaatiossa ja muilla nisäkkäillä tällaista etua ei kuitenkaan ole, sillä genomit ovat isoja, geenien välillä on paljon vuorovaikutusta, ja lisääntymiskohinaa on paljon. Näin ollen meillä on valinnan ulottumattomissa olevia nukleotideja paljon enemmän kuin mikrobeilla.
Merkitykseltään vähäiset mutaatiot
Heti lähes neutraaleja mutaatioita suurempivaikutukselliset mutaatiot ovat merkitykseltään vähäisiä mutaatioita. Niillä on havaittavissa oleva vaikutus, vaikka se onkin pieni. Jalostajilla on vaikeuksia nähdä näitä, mutta luonnonvalinta usein ”huomaa” ne, jos populaatiolla on riittävästi kokoa ja aikaa on riittävästi käytettävissä. Tällä on kuitenkin rajoituksensa, sillä näiden mutaatioiden vaikutus on pieni ja muiden tekijöiden vaikutus voi olla suurta. On kuin yrittäisi nähdä myrskyävästä merestä sinne heitetyn kiven aiheuttamat aallot.
Merkitykseltään vähäisten mutaatioiden vaikutuksen yli ajaa mm.
- Populaation suuri satunnainen kuolleisuus
- Tappavat ja lähes tappavat mutaatiot
- Suurin osa mutaatioista on peittyviä
- Geenien välinen vuorovaikutus (epistasia)
- Pienissä populaatioissa seksuaalisen rekombinaation satunnaisuus
Nämä tekijät aiheuttavat geneettistä ajautumista. Vaikka luonnonvalinta pystyykin poistamaan useita tällaisia merkitykseltään vähäisiä mutaatioita, tulee näistä mutaatioista kohinaa toisilleen, kun niiden määrä kasvaa. Valinnan teho tippuu nollaan hyvin nopeasti, kun valittavana olevien mutaatioiden määrä kasvaa. Kimura35 viittaa tähän valintainterferenssiksi kutsuttuun ilmiöön, ja esittää (s. 30), että vain 138 nukleotidipaikkaa on yhtä aikaa valittavissa, vaikka populaatiosta voitaisiin poistaa 50% joka sukupolvessa ja yhtä ominaisuutta vastaan valitaan heikosti (s = 0,01).
Useampien satojen mutaatioiden yhtäaikainen valintayritys pysäyttää valinnan vauhdin täysin ja näin varsin rajoitettuunkin määrään mutaatiota vaikuttamiseen menee satoja tuhansia sukupolvia. Tämä rajoitus tekee genominlaajuisen valinnan mahdottomaksi.
Lisääntymisen estäminen
Mutantteja ei voida järjestelmällisesti estää lisääntymästä logistisista syistä, mikä tuottaa ongelman valinnan kannalta.33, 45, 43 Toivottu ominaisuus on tyypillisesti yksilössä, jossa on myös jokin ei-toivottu ominaisuus, eikä näitä ominaisuuksia voida erottaa toisistaan. Koska mutaatioista suurin osa on haitallisia, päätyy näitä väkisin enemmän seuraavalle sukupolvelle, kun valitaan jonkin hyödyllisen puolesta.
Jos otetaan samasta populaatiosta kaksi yksilöä, joilla kummallakin on omat 10 000 mutaatiota perimässään. Näin ollen kahden yksilön välillä on 20 000 eroa, ja kummallakin on 10 000 hyvää ja saman verran huonoja nukleotidipaikkoja toiseen yksilöön nähden. Keskiarvoinen ero on pieni ja kelpoisuuden kannalta sen ratkaisevat harvat merkittävät mutaatiot, joihin valinta kohdistuu samalla kun enemmistö mutaatioista jää valinnan ulottumattomiin. Näin ei voida estää mutaatioiden kertymistä, vaikka lisääntymisen suhteen voitaisiinkin ylläpitää täydellistä kontrollia.
Fyysinen kytkentä
Koko genomi koostuu suurista yhteenlinkittyneistä ryhmistä46. Jokainen tällainen blokki sisältää vahingollisia muutoksia, vaikka se sisältäisi jonkun merkittävän hyödyllisenkin mutaation. Ryhmän kokonaisvaikutus kelpoisuuteen on siis vahingollinen, vieläpä, kun kuhunkin tällaiseen ryhmään ehtii kertyä tuhansia vahingollisia mutaatioita ennen ensimmäistäkään hyödyllistä.
Vuonna 2006 julkaistun tutkimuksen47 mukaan ihmisen olisi pitänyt kuolla sukupuuttoon evolutiivisella aikajanalla. Tutkimuksessa käsiteltiin kuitenkin vain mitokondrion DNA:ta, ja se on vain yksi ihmisen 100 000 – 200 000:sta yhdistyneestä ryhmästä. Näin ollen tutkimus aliarvioi yhteenliittymien ongelman jopa 200 000-kertaisesti.
Luku 6: Kohina
Jos haluaa vastaanottaa signaalin, se täytyy erottaa kohinasta tai muista häiriöistä, eli signaalin ja kohinan välisen suhteen pitää olla riittävän suuri. Genetiikassa signaalin ja kohinan välistä suhdetta kuvataan periytyvyydellä. Jos ominaisuuden periytyvyys on suuri, se on valinnan ulottuvilla, mutta jos periytyvyys on pieni, suurin osa ominaisuuteen vaikuttavista tekijöistä on ympäristötekijöitä. Moniin ominaisuuksiin, kuten pituuteen, vaikuttavat sekä perintötekijät, että ympäristötekijät. Toisaalta esimerkiksi veriryhmä periytyy täysin geenien perusteella (periytyvyys 1), ja tatuoinnit eivät periydy ollenkaan (periytyvyys 0).
Merila ja Shelton48 ja Kruuk ym.49 ovat osoittaneet, että luonnollisissa populaatiossa kelpoisuuden periytyvyys on tyypillisesti periytymättömien ympäristötekijöiden tuottaman kohinan takia liian pientä mitattavaksi. Aiemmin oletettiin, että tämä lähes olematon periytyvyys johtuu niin voimakkaasta valinnasta, että geneettistä monimuotoisuutta ei jäänyt jäljelle. Nykyään kuitenkin tiedämme mutaationopeuksien olevan suuria, jolloin geneettistä vaihtelua näissä populaatioissa kyllä on.
Kelpoisuuden vähäinen periytyvyys tarkoittaa, että suurin osa kelpoisuuteen vaikuttavista tekijöistä on periytymättömiä ympäristötekijöitä, jolloin huonojen yksilöiden poistaminen vaikuttaa vain hyvin vähän huonojen geenien poistamiseen. Esimerkiksi puun siemenistä osa putoaa hedelmälliseen maaperään, mutta suurin osa siemenistä tippuu kuitenkin jollain tavalla sopimattomaan, esimerkiksi liian märkään tai kuivaan, maaperään. Lähes kaikki tällaisesta kelpoisimman valinnasta onkin onnekkaimman valintaa, jolla ei ole yhteyttä geneettisesti parhaan valintaan.
Usein on arvioitu, että noin puolet kaikista fenotyypin muutoksista perustuu ympäristötekijöihin. Esimerkiksi kahdesta kasvista pellolla paremmin kasvavan kasvin kasvuolosuhteet ovat paremmat. Noin neljännes kaikista fenotyypin muutoksista taas perustuu genotyypin ja ympäristötekijöiden yhteisvaikutukseen. Esimerkiksi jos peltoa kastellaan, voi toiselle kasville lisävesi olla hyväksi, toiselle huonoksi, genomista riippuen. Nämä muuntelun tyypit vähentävät signaalin ja kohinan välistä suhdetta ja häiritsevät valintaa.
Kolmas signaalin ja kohinan välistä suhdetta heikentävä tekijä on periytymätön geneettinen muuntelu, kuten epigeneettiset, epistaattiset, vallitsevuus- ja syklisen valinnan vaikutukset. Näiden lisäksi on sitten niin sanottu lisäävä geneettinen vaihtelu, joka tarkoittaa sitä, että jokin ominaisuuden tai nukleotidin muutos on selvästi vaihtoehtoaan parempi. Kimuran35s. 30-31 mukaan tällainen muuntelu käsittää noin 0,4% kelpoisuuteen vaikuttavasta muuntelusta. Jos Kimura on tässä oikeassa, se tarkoittaa, että 99,6% fenotyypin kelpoisuuteen vaikuttavasta valinnasta menee täysin hukkaan.
Jos vielä populaatio lisääntyy hitaasti, kuten ihmiset, todella tehokkaaseen valintaan voidaan käyttää vain hyvin pientä osaa populaatiosta. Jos esimerkiksi 16,7% populaatiosta voidaan valinnallisesti poistaa ja 0,04% valinnasta todella vaikuttaa, on toimivan valinnan ulottuvissa vain 0,07% populaatiosta – eli alle yksi tuhannesta.
Jos tarkastellaan yksittäisen nukleotidimuutoksen periytyvyyttä, havaitaan, että sen kelpoisuutta parantava arvo (signaali) on häviävän pieni samalla, kuin kaiken periytymättömän muuntelun ja kaikkien muiden nukleotidipaikkojen yhteisvaikutus (kohina) on tähtitieteellinen. Tästä syystä suurin osa mutaatioista on valinnan ulottumattomissa.
Aiempien lisäksi kohinaa tuottaa todennäköisyysvalinta. Siinä, missä jalostajat pisteyttävät jalostettavat eliönsä erilaisten parametrien, kuten elinvoiman ja tuottoisuuden mukaan ja sitten määrittelevät rajan, jota heikommat karsitaan täysin ja jota paremmat saavat lisääntyä. Luonnossa näin ei kuitenkaan tapahdu, sillä muutokset saavat aikaan vain eroja lisääntymistodennäköisyyteen ja sattumalta huonommatkin yksilöt saavat lisääntyä.
Luonnossa valinta onkin satunnaista onnekkaimman valintaa, ja siitä puuttuu lähes täysin fenotyypin mukainen järjestelmällinen valinta. Esimerkiksi sammakon lisääntyessä munista ison osan syövät kalat, sammakonpoikasista linnut, vene liiskaa osan ja monet tippuvat vesiputouksesta. Täysikasvuisista sammakoista osa kaivautuu rantatörmään, joka ruoppausoperaation yhteydessä poistetaan. Lisääntymisen epäonnistumisesta jopa puolet johtuu vastaavista fenotyypistä riippumattomista tekijöistä.
Kolmas kohinaa tuottava tekijää kutsutaan sukusolujen otannaksi. Se tarkoittaa sitä, että kaikki tarjolla olevat sukusolut eivät hedelmöity, vaan vain jotkin niistä. Tämä johtaa siihen, että ominaisuuksien periytyminen vaihtelee kuten kolikonheiton tulokset. Jos kolikkoa heitetään monia kertoja, tulokseksi saadaan hyvin tarkasti puolet kruunia ja puolet klaavoja, mutta jos kolikkoa heitetään vain muutama kerta, suhde on hyvin usein jotain ihan muuta. Tästä seuraa geneettinen ajautuminen, joka pienissä populaatioissa on hyvin merkittävä, mutta isommissa selvästi pienempi. Tällaisen kohinan voi siis saada pois kuvittelemalla mahdollisimman suuren populaation ja toisaalta kohinan voi lisätä äärimmilleen olettamalla pienen populaation.
Isotkaan populaatiot eivät kuitenkaan ole vapaita kohinan vaikutuksista, sillä fenotyyppiin ja lisääntymismenestykseen vaikuttava kohina vaikuttaa niihinkin. Hyvin suurissa populaatioissa kohina myös vahvistuu alapopulaatioiden oman sukusolujen otannan ja monien erilaisten ympäristöolosuhteiden takia. Siinä missä jalostajat ovat voineet käyttää tilastollisia menetelmiä ja karsivaa valintaa valitakseen rajattuja määriä ominaisuuksia, ei luonnonvalinnalla ole käytössään mitään niistä.
Kohinan seurauksena valinnan ulottumattomissa olevien mutaatioiden määrä lisääntyy merkittävästi, valintakynnysongelma, jossa usean yhtäaikaisen ominaisuuden valinta johtaa valinnan pysähtymiseen, pahenee ja yhä suurempi osa valinnasta menee hukkaan.
Punainen vaunu -analogiassa kohina tarkoittaisi työntekijöiden virheitä, jolloin laadunvalvoja haaskaisi voimavaransa virheisiin, jotka eivät edes seuraa kokoamisohjekirjan virheistä. Prinsessan tapauksessa patjoissa olisi lukuisia herneen kokoisia möykkyjä, ja oppikirja-analogiassa oppikirjat vaihdettaisiin kesken lukukautta. Herneen kokoiset möykyt patjoissa tekisivät patjojen alla olevien muutosten havaitsemisesta paljon vaikeampaa ja oppikirjojen vaihtaminen vähentäisi edelleen olemattoman pientä korrelaatiota oppikirjojen kirjoitusvirheiden ja oppimistulosten välillä.
Luku 7: Keinotekoiset pelastusmekanismit
Koska tutkimuksissa on havaittu, että mutaatioita kertyy perimään paljon, on tämä mutaatioiden kertymisnopeusongelma täytynyt ratkaista joillain tavalla. Yksi selitystapa on ollut roska-DNA, mutta koska tuoreemmat tutkimukset ovat paljastaneet, että roska-dna:ta ei olekaan – ainakaan yhtään niin paljoa, kuin on väitetty – on keksitty kaksi mekanismia mutaatioiden kertymisongelman ratkaisemiseksi: mutaatioiden määrään perustuva mekanismi ja synergistisen epistasian mekanismi.
Mutaatioiden määrään perustuvan mekanismin mukaan joillain yksilöillä on sattumalta enemmän mutaatioita kuin toisilla, ja kohdistamalla valinta näihin yksilöihin, päästään eroon suuremmasta määrästä mutaatioita pienemmällä valinnan hinnalla.
Mekanismin ongelmana on kuitenkin se, että mutaatioiden määrää olennaisempaa on mutaatioiden vaikutus: mitä pahempia mutaatioita, sen huonompi. Yhden merkittävän mutaation vaikutus voi olla suurempi kuin tuhannen vähemmän vaikuttavan mutaation. Lisäksi esimerkiksi eri ihmisten mutaatioiden määrien väliset erot ovat vähäisiä keskiarvon molemmin puolin ja johtuvat geneettisestä ajautumisesta, joten yksilöiden välillä ei ole merkittäviä eroja.
Mutaatioiden määrään perustuvan mekanismin simulaatiokokeissa50 on todettu, että mekanismi toimii vain, jos valinta on tiukan karsivaa, ja kaikkien mutaatioiden vaikutus on sama. Tällaiset olosuhteet ovat kuitenkin erittäin keinotekoiset, ja todellisuudessa vallitsevissa olosuhteissa, joissa vaikuttaa todennäköisyysvalinta ja mutaatioiden vaikutukset ovat erilaisia, populaatiot rappeutuvat sukupuuttoon asti.
Synergistinen epistasia tarkoittaa sitä, että muiden vahingollisten mutaatioiden vaikutus vahvistaa toisten vahingollisten mutaatioiden vaikutusta. Näin kun mutaatioita kertyy enemmän, niiden vaikutus on huomattavampi ja näin ne tulevat ”valinnan näkyviin”. Todellisuudessa populaatiogeneetikot ovat kuitenkin yksimielisiä siitä, että tällainen on harvinaista, ja yleensä mutaatioiden yhteisvaikutus on joko lisäävä tai moninkertaistava.
Jos synergististä epistasia olisi yleisempi, se tarkoittaisi, että eliöt rappeutuvat nopeammin kuin muuten, kuten on osoitettu simulaatiokokeilla.51 Yleisesti on myös tiedossa, että geenit vaikuttavat toisinsa (epistasia) ja tämä haittaa merkittävästi valintaa. Molemmat mekanismit toimivat siis vain erittäin keinotekoisissa olosuhteissa, eivätkä todellisessa maailmassa pysty estämään mutaatioiden kertymistä.
Luku 8: Rodunjalostus ja kloonaus?
Rodunjalostus ei voi pelastaa genomia, sillä valinnalla voidaan valita vain hyvin rajattua määrää ominaisuuksia, samalla, kun lähes neutraalit mutaatiot kertyvät edelleen perimään. Lisäksi paremmuus ja huonommuus ovat hyvin heikosti periytyviä ominaisuuksia, joten niiden valinta kohdistuisi lähes kokonaan genetiikasta riippumattomiin tekijöihin. Lisäksi ihmisen lisääntymisen säännöstely olisi niin poliittisesti kuin sosiaalisestikin mahdotonta.
Kloonaaminenkaan ei pelasta genomia, sillä Müllerin säppipyörä6, 52 pyörii vain yhteen suuntaan. Jokainen kloonattu yksilö on aiempaa mutatoituneempi, eikä tätä mutaatioiden kertymistä estämään ole mitään mekanismia. Sikäli, kun kloonattuja kasveja vielä on elossa, se johtuu siitä, ettei vielä ole kulunut riittävästi aikaa niiden päätymiseksi sukupuuttoon.
Eläimillä kloonaamisella ei saavuteta kasvien tavoin edes lyhyen aikavälin hyötyjä, sillä kloonit ovat vanhentuneet jo ennenaikaisesti. Kehon soluihin tulee paljon enemmän mutaatioita kuin sukusoluihin, jotka jakautuvat mahdollisimman vähän. Näin ollen kloonia varten otettu solu on jo vanhentunut selvästi etukäteen ja klooni on alkuperäistä yksilöä rappeutuneempi. Kloonaamalla nähtäisiinkin vähän siitä, mihin ihmiskunta on menossa: suunta on alaspäin, ei ylöspäin.
Luku 9: Voiko luonnonvalinta luoda?
Jos jätetään kaikki vahingolliset mutaatiot huomiotta, voisiko siinä tapauksessa mutaatioiden ja luonnonvalinnan kautta syntyä uusi toimiva geeni? Yksi toimiva geeni kuvaa kirjan lukua tai toiminnallista ohjelmaa ja on vähintään 1000 kirjainta pitkä. Tällaisen tuhannen kirjaimen mittaisen geenin synnyssä on kuitenkin ongelmansa.
1. Ensimmäisen halutun mutaation määrittely.
Yksittäisellä nukleotidilla ei ole merkitystä yksinään, vaan vain ympäröivien nukleotidien kontekstissa samalla tavalla kuin kirjoitetussa tekstissä yksittäisellä kirjaimella ei ole yksinään mitään erityistä merkitystä. Kokoamisohjeanalogiasta näkyy myös, että nykyisessä kontekstissa mahdolliset hyödylliset muutokset ovat hyvin vähäisiä. Uutta geeniä luotaessa on kuitenkin tarkoitus muuttaa laajemmin kontekstia. Samalla kun yksittäistä nukleotidia pyrittäisiin kehittämään ympäröivien nukleotidien kontekstissa, tuhotaan tätä kontekstia kyseisen nukleotidin ympäristössä, sillä muitakin nukleotideja täytyy muuttaa.
2. Ensimmäistä mutaatiota odotellessa
Jos kuitenkin oletamme, että ensimmäinen haluttu mutaatio on selvillä, kuinka kauan sitä täytyisi odottaa? Yksittäisen nukleotidipaikan mutaationopeus on äärimmäisen pieni, karkeasti arvioituna yhteen tiettyyn nukleotidipaikkaan ihmisellä tulee yksi muutos 30 miljoonaa yksilöä kohden. Jos myös oletamme ihmisen evoluution tapahtuneen noin 10 000 yksilön populaatiossa, tarkoittaisi tämä yhden nukleotidipaikan muuttumisen vievän 3000 sukupolvea eli noin 60 000 vuotta. Kyseinen nukleotidi voi kuitenkin vaihtua myös kahteen muuhun nukleotidiin halutuun nähden, joten käytännössä aika tarvittaisiin kolme kertaa kauemmin – 180 000 vuotta.
Mutaation tapahtuminen ei kuitenkaan vielä riitä, vaan sen pitää myös vakiintua populaatioon. Satunnaisen geneettisen ajautumisen takia ylivoimainen enemmistö mutaatioista ei vakiinnu populaatioon. Mutaation tulisi olla vallitseva ja tuottaa merkittävä etu, jotta valinta voisi valita sen puolesta, mutta tiedämme lähes kaikkien hyödyllisten mutaatioiden olevan lähes neutraaleja ja peittyviä.
Jos valinta ei ole tehokasta, 10 000 yksilön populaatiossa olisi sattumalta vain yksi mahdollisuus 20 000 tuhannesta, että mutaatio vakiintuu populaatioon, sillä näin paljon on muuttumattomia nukleotidipaikkoja populaatiossa. Sanfordin ja hänen kollegojensa simulaatioiden perusteella 99% lievästi hyödyllisistä mutaatioista poistuu populaatiosta satunnaisen ajautumisen takia.
Tavanomaisen hyödyllisen muutoksen tulisi siis tapahtua noin sata kertaa, ennen kuin se vakiintuu populaatioon. Tämä tarkoittaa, että aikaa tarvittaisiinkin 180 000 * 100 vuotta, eli 18 miljoonaa vuotta ensimmäisen halutun mutaation vakiinnuttamiseksi populaatioon. Oletus kuitenkin on, että ihminen on eronnut edellisestä simpanssin kanssa yhteisestä kantamuodosta kuusi miljoonaa vuotta sitten, joten aikaa ei ole ollut edes ensimmäisen halutun mutaation vakiinnuttamiseksi populaatioon.
3. Muita mutaatioita odotellessa
Kun ensimmäinen mutaatio on saatu vakiinnutettua populaatioon, täytyy myös loppujen 1000 muutoksen vakiintua populaatioon. Jokaisella näistä mutaatioista se vie noin 18 miljoonaa vuotta, eli kokonaisuudessaan 18 miljardia vuotta. Tämä on jo enemmän kuin mitä alkuräjähdyksestä on aikaa. Käytännössä myös useiden nukleotidien pitäisi muuttua kerralla ja vakiintua populaatioon, sillä yksittäinen nukleotidi ei vielä itsessään sisällä informaatiota. Jos valinta alkaa toimia esimerkiksi 12 nukleotidipaikan muututtua halutuiksi, niin odotusaika kasvaa tässä 10 000 yksilön ihmispopulaatiossa biljooniin vuosiin.
4. Haldanen dilemman odotus
Mutaation tulee vakiinnuttuaan myös levitä koko populaatioon. 10 000 yksilön populaatiossa on 20 000 nukleotidipaikkaa, johon sen pitäisi levitä. Jos oletetaan hyvin voimakas valinta, joka levittää tätä mutaatiota 10% lisää populaatioon joka sukupolvi, menee 105 sukupolvea (1,1^105 = 20 000), että se on levinnyt populaatioon. Tällaista valintaa ei kuitenkaan yleensä ole. Jos mutaatio on peittyvä, sen leviäminen populaatioon vie vähintään 100 000 sukupolvea, vaikka se parantaisi sopeutuvuutta prosentilla53.
Tätä ongelmaa kutsutaan tyypillisesti Haldanen dilemmaksi, sillä Haldanen33 mukaan menisi keskimäärin 300 sukupolvea eli 6000 vuotta saada yksi uusi mutaatio leviämään populaatioon. Tämä arvio perustuu hänen arvioonsa siitä, mikä on vallitsevien ja peittyvien muutosten suhde. Tämän laskelman perusteella voisi siis 1000 muutosta levitä populaatioon oletettuna 6 miljoonan vuoden aikana, mutta nämä olisivat vain yksittäisiä toisistaan irrallisia hyödyllisiä mutaatioita, ei yhden uuden geenin luomiseen tarvittavia toisiinsa linkittyneitä muutoksia.
Valinta voisikin vakiinnuttaa vain 1000 yksittäistä muutosta oletetun ihmisen kehityshistorian aikana, eikä useampia muutoksia voida valintainterferenssin takia valita yhtä aikaa. Tämä on erittäin vähän informaatiota – vähemmän kuin tavallisella verkkosivulla – eikä se riitä erottamaan ihmistä simpanssista. Ongelman ovat itsenäisesti todenneet Crow ja Kimura54, ReMine36, 37 ja Rupe ja Sanford55.
5. Rekombinaatiota odotellessa
Voitaisiin ajatella, että sopivat mutaatiot tapahtuvat populaation eri yksilöissä ja sitten rekombinaatiossa ne yhdistyisivät, jolloin kehitys tapahtuisi nopeammin. Tähän liittyy kuitenkin kaksi ongelmaa:
Ensinnäkin ihmisen geeniperimä koostuu suurista 20 000 – 40 000 nukleotidin yksiköistä, joiden sisällä ei ole tapahtunut rekombinaatiota nykyihmisen olemassaoloaikana.56, 46 Näin ollen rekombinaatiota ei tapahdu yksittäisten nukleotidien tasolla. Toiseksi todennäköisyys sille, että näin rekombinaatiolla sekoittamalla saataisiin haluamamme 1000 nukleotidin geeni, on äärimmäisen pieni. Lisäksi tällainen rekombinaatio hajottaisi ketjuja nopeammin kuin synnyttäisi niitä, joten aikaa tarvittaisiin enemmän.
6. Loputtomat kelpoisuuslaaksot
Kun uusi geeni on kehittymässä, se tuottaa haittaa lajin menestykselle, eli laskee kelpoisuutta. Tästä evoluutioteoreetikot ovat yksimielisiä. Uuden geenin kehitys vaatii monia kokeiluja ja nämä ovat paljolti vahingollisia. Lisäksi puolivalmis geeni ei ole vielä hyödyllinen. Näin ollen populaatiot päätyvät kelpoisuuslaaksoihin, jossa kehittyvä laji tulee huonommaksi ennen kuin se voi kehittyä.
Kun yhdistetään hyvien mutaatioiden harvinaisuus Haldanen dilemmaan, havaitaan, että näistä kelpoisuuslaaksoista tulee erittäin pitkiä ja syviä, erityisesti kun evolutiivisia kehitysaskelia täytyy ottaa koko ajan. Kelpoisuuslaaksojen pitäisi kuitenkin olla lyhyitä ja matalia, jotta populaatiot eivät päätyisi sukupuuttoon. Kun tarkastellaan tilannetta koko perimän tasolla, on menossa lukuisia hyvin syviä kelpoisuuslaaksoja koko ajan, mikä osoittaa evolutiivista kehitystä vastaan.
7. Monirajoitettu DNA
Kuvitellaan teksti, jossa on yksi viesti, kun se luetaan etuperin, toinen viesti, kun se luetaan takaperin, kolmas viesti, kun tekstistä luetaan vain joka toinen kirjain ja neljäs viesti, kun tekstin purkamiseen käytetään monimutkaista salauksenpurkualgoritmia. Tällainen viesti on monirajoitettu ja monitoiminnallinen, eikä mikään kirjoitusvirhe voi sitä parantaa, vain tuhota.
DNA:n monitoiminnallisuudesta on runsaasti näyttöä. Suurin osa ihmisen DNA-sekvensseistä koodaa kahta eri RNA:ta, joita luetaan molempiin suuntiin.57 Monet DNA-sekvenssit koodaavat eri toimintoja riippuen siitä, mistä lukeminen aloitetaan, tai lähetti-RNAn silmukoitumisen perusteella. Lisäksi todennäköisesti kaikki sekvenssit DNA-koodissa vaikuttavat DNA:n kolmiulotteiseen rakenteeseen.
8. Palautumaton monimutkaisuus
Pieni leluvaunu -analogiassa voitaisiin ajatella pienen kehitysaskeleen olevan punaisen leluvaunun muuttuminen siniseksi kolmipyöräiseksi. Värin koodin muutoksen aikaansaava kirjoitusvirhe on helppo kuvitella – kunhan sininen maali on saatavissa ja koodattu. Myös yksi pyörä voi tippua pois, mutta kolmepyöräinen leluvaunu ei ole kolmipyörä, vaan rikkoutunut leluvaunu. Käytännössä muutos leluvaunusta kolmipyörään vaatisi koko kokoamisohjeen perusteellista uudelleenkirjoittamista.
Suurinta osaa osista pitää muuttaa radikaalisti, eikä toimivia välimuotoja ole, joten analogian laadunvalvoja valitsisi haluamiamme muutoksia vastaan, koska se ei näe tulevaisuutta. Oikean kirjoitusvirhekokonaisuuden tulisi siis sattumalta ilmestyä kerralla, eikä näin koskaan tapahdu – muutokset voivat syntyä vain älykkään toiminnan tuloksena.
Yksinkertaisimmassakin proteiinissa on paljon enemmän monimutkaisuutta kuin leluvaunussa, joten palautumatonta monimutkaisuutta on myös paljon enemmän. Proteiinien perustana olevassa geneettisessä koodissa monimutkaisuutta on vieläkin enemmän. Pelkästään DNA:n design on hämmästyttävä, puhumattakaan muusta molekyylikoneistosta.
Yksittäinen geeni käsittää kaikkine säätelyosineen ja arkkitehtuuriin liittyvine elementteineen noin 50 000 osaa – enemmän kuin monessa nykyaikaisessa autossa. Kuitenkin tämä on vain hyvin pieni osa koko solun monimutkaisuutta, jossa on palautumatonta monimutkaisuutta palautumattoman monimutkaisuuden päällä. Voimme tehdä hyvin pieniä keinotekoisia geenejä, mutta emme täysin uutta geeniä aiemmin tuntemattomalle proteiinille ja saada sitä toimimaan solussa. Kuinka sitten sattumanvaraiset muutokset yhdistettynä hyvin rajalliseen määrään lisääntymissuodattumista?
9. Melkein kaikki hyödylliset mutaatiot ovat lähes neutraaleja
Lähes neutraalien mutaatioiden näkymättömyys valinnalle ja siitä seuraavat ongelmat on geneetikkojen parissa laajalti hyväksytty, mutta ei sen sijaan sitä, että hyödyllisillä mutaatioilla tämä lähes neutraaliuden ongelma on vielä pahempi. Toivomallamme geenillä on joitain vaikutukseltaan merkittäviä nukleotideja, esimerkiksi entsyymin aktiivisen keskuksen määräävät nukleotidit. Nämäkin ovat turhia ja valinnan ulottumattomissa ilman suuria määriä itsessään hyvin vähäisesti merkittäviä nukleotideja. Näitä nukleotideja ei voida asettaa eikä pitää paikallaan valinnan kautta, joten ensimmäisestäkään halutusta nukleotidista ei saada kiinni.
10. Vahingolliset mutaatiot takaisin kuvaan
Rekombinaatio toimii ennemmin geenien, kuin nukleotidien tasolla, sillä ihmisen genomissa näkyy suuria DNA-ryhmittymiä, joissa ei ole tapahtunut nykyihmisen historian aikana rekombinaatiota56, 46. Tällaiset dna-ryhmittymät voivat joutua Müllerin säppipyörään,6, 47 joka tarkoittaa, ettei haitallisia mutaatioita voida erottaa hyödyllisistä. Koska haitallisia mutaatioita on valtavasti enemmän kuin hyödyllisiä, kaikki tällaiset osat DNA:ta rappeutuvat. Vaikka joitakin hyödyllisiä mutaatioita saataisiinkin vakiinnutettua, haitalliset muutokset tekevät niistäkin selvää loppuja hyödyllisiä odotellessa.
Valinnalla on myös hintansa. Hyödyllisten mutaatioiden valintaan voidaan kohdistaa valintaa vain, jos kaikki muut lisääntymishinnat on katettu, mukaan lukien haitallisten mutaatioiden poistamisesta johtuva hinta. Lyhyellä aikavälillä voidaan kohdistaa valintaa myös hyödyllisiin mutaatioihin, pitkän aikavälin kustannuksella.
Mutaatiot eivät myöskään ole täysin satunnaisia, sillä osa genomista mutatoituu paljon useammin kuin toiset osat. Useimmin mutatoituvissa niin sanotuissa kuumissa pisteissä haluamamme mutaatio tulee nopeasti, mutta sillä välin kun odotetaan hitaasti mutatoituvissa paikoissa tapahtuvia muutoksia, ehtivät näissä kuumissa pisteissä nukleotidit vaihtua jo pois haluamistamme. Lisäksi monilla nukleotidipaikoilla on taipumus mutatoitua johonkin tiettyyn nukleotidiin, kuten T. Tämä auttaa silloin, kun T on haluttu, mutta hidastaa, kun A, C tai G on haluttu. Tämä edelleen hidastaa kehittävää valintaa.
Vuonna 2002 julkaistun tutkimuksen58 mukaan ihminen ja simpanssi eroavat noin 150 miljoonassa nukleotidipaikassa 40 miljoonan mutaation seurauksena. Näin ollen sekä simpanssi että ihminen ovat saaneet noin 20 miljoonaa mutaatiota kehityslinjallaan, mutta vain noin 1000 näistä on voitu valinnalla levittää populaatioon – loput ovat kertyneet satunnaisen ajautumisen kautta. Vain noin 1000 hyödyllistä muutosta ja noin 20 miljoonaa vahingollista: tämä ei olisi vain tehnyt meistä kantamuotojamme huonompia, vaan olisi tappanut meidät sukupuuttoon.
Johtopäätökset
Geenien täytyy olla suunniteltuja, eivätkä ne ole voineet kehittyä. Suurimman osan käyttökelpoisesta vaihtelusta täytyykin olla suunniteltua. Sopeuttavaa valintaa kuitenkin tapahtuu, mutta se ei johdu satunnaisten mutaatioiden tuottamasta uudesta informaatiosta, vaan jo olemassa olevan vaihtelupotentiaalin puitteissa tapahtuvasta hienosäädöstä. Kyse on alunperinkin segregoitumaan ja rekombinoitumaan suunniteltujen rakenteiden segregoitumisesta ja rekombinoitumisesta. Tällä tavoin syntyy valtava määrä muuntelupotentiaalia, sillä jo yhden ihmisparin jälkeläisille voi tulla valtava määrä erilaisia muunnoksia. Tällaisen muuntelun kanssa luonnonvalintakin toimii tehokkaimmin. Samoin kuin järjestetyssä korttipakassa informaatio on suurimmillaan alussa ja monimuotoisuus useamman pelikierroksen jälkeen, ei ihmiselläkään alun jälkeen tarvita uutta informaatiota.
Luku 10: Vastaako laskeva käyrä todellisuutta?
Näkemys alati etenevästä evoluutiosta on niin syvään juurtunut ajatteluumme, että on vaikea uskoa näin nopeaan geneettiseen rappeutumiseen suuresta mutaatiovauhdista, valinnan hinnasta, lähes neutraaleista mutaatioista, valintakynnyksestä ja valintainterferenssistä johtuen. Johtavat populaatiogeneetikot ovat kuitenkin aiempien vuosikymmenten aikana tuoneet esiin vakavia epäilyksiä siitä, että näistä ongelmista olisi mahdollista selvitä.
Sanford kirjoittaa tutkineensa vuosikymmenen ajan huolellisesti mutaation ja valinnan prosesseja, ja geneettistä entropiaa ehkä enemmän kuin kukaan muu. Lisäksi hän on kehittänyt numeerisen simulaatio-ohjelmiston, jolla on mahdollista simuloida realistisesti mutaatioiden ja valinnan vaikutusta, ottaen huomioon kaikki oleelliset tekijät samanaikaisesti. Simulaatioiden lisäksi on olemassa kokeellista näyttöä RNA-virusten, kuten influenssavirusten geneettisestä entropiasta. Näiden taudinaiheutuskyky laskee nopeasti mutaatioiden kertymisestä johtuen.59, 60
Luonnonvalintaa on pidetty geneettisen rappeutumisen vastavoimana, mutta erityisesti isoissa genomeissa mutaatioiden aiheuttama rappeutuminen on niin suurta, ettei valinta voi sitä kumota. Näin ollen nämä genomit väistämättä tuhoutuvat, ellei ole jotain vielä tuntematonta valinnasta riippumatonta vastavoimaa. Tästä on kyse geneettisessä entropiassa, joka on johtopäätös mutaatioiden ja valinnan todellisten toimintatapojen huolellisesta analyysistä.
Mutaation ja valinnan kyvyttömyys pysäyttää genomin rappeutumista osoittaa evoluutioteorian täysin virheelliseksi – ei vain epäuskottavaksi tai epätodennäköiseksi. Se on vain perusteeton hypoteesi. Tämä johtuu jo siitä, että valinta kohdistuu yksilöön, eikä genomiin. Se ei siis voi estää molekyylitasolla tapahtuvien muutosten aiheuttamaa äärimmäisen monimutkaisten järjestelmien tuhoutumista. On kuin yrittäisi korjata tietokonetta vasaralla.
Kirjan alussa käytettiin analogiaa pienestä leluvaunusta, jonka kokoamisohjeita kopioi lähes sokea kirjoitusvirheitä tekevä kirjuri-robotti, ja kuviteltiin, kuinka lähes sokea tuomari-robotti valvoi laatua. Älyllistä ohjausta ei ole. Kysymys on, tuleeko tällaisella tuotantolinjalla autoista avaruusaluksia. Varsin selvää on, ettei tule, vaan vain huonompia autoja. Tuomari-robotti voi vain hidastaa autojen huononemista. Entä jos annettaisiin äärettömästi aikaa? Se tarkoittaisi vain täyttä varmuutta, että tuotantolinjalta ei lopulta tule mitään toimivaa ulos.
Intuitiivisesti on selvää, ettei kirjoitusvirheiden kautta tuotantolinjan autoihin tule rakettimoottoria. Vaikka tuomari-robotti näkisikin tällaisen auton, se valitsisi sen pois, sillä rakettimoottori ei tee autosta parempaa autoa. Tuomari-robotti kun ei näe tulevaisuutta, eikä sillä ole käsitystä avaruusaluksesta, ainoa, mitä se voisi tehdä avaruusaluksen aikaansaamiseksi, on valita parempia autoja – ja tämä on paradoksaalista.
Edes se, että tuomari-robotti voisi alkaa valita enemmän avaruusalusta näyttäviä autoja, ei mahdollista autojen kehitystä avaruusaluksiksi, sillä tarvittavien kirjoitusvirheiden määrä on niin valtava, ettei aika tehtaalla millään riitä. Lisäksi avaruusaluksen kaltaiset autot ovat toimimattomia tuotteita, ne eivät toimi sen paremmin autoina kuin avaruusaluksina. Tällaisen tehtaan vääjäämättömänä kohtalona olisi konkurssi.
Yksilön vanheneminen on samankaltainen prosessi lajin vanhenemiseen nähden. Molemmat johtuvat mutaatioiden kertymisestä. Ihmisen eliniänodote on noin 70 vuotta maksimin ollessa noin 120 vuotta. Serkusten mennessä naimisiin eliniänodote on pienempi. Tämä johtuu peittyvien muutosten esiintulosta. Jos ne kaikki tulisivat esiin nyt, kuolisimme heti sukupuuttoon.
Raamatussa kuvataan ajanjakso, jonka aikana ihmiset elivät yli 900-vuotiaiksi ja sisäsiittoisuus oli vaaratonta. Tämä tuntuu aivan järjettömältä, mutta toisaalta ymmärryksemme on rajallista. Emme esimerkiksi tiedä, miksi nisäkkäiden elinikä on tyypillisesti maksimissaan alle 20 vuotta ja ihmisellä noin 120 vuotta. Simpanssillakin maksimielinikä jää alle puoleen 120 vuodesta. Ikääntymisen tiedetään johtuvan mutaatioista, joten jos alun perin mutaatioita ei ollut, ovat yli 900-vuoden eliniät loogisia.
Nooasta seuraavien sukupolvien aikana näkyy myös, kuinka eliniät laskevat eksponentiaalisesti.61 Joko Raamattuun on kuvattu todellisia elinikiä tai sitten Raamatun kirjoittajilla on ollut huomattava tarve esittää ekspotentiaalinen rappeutumiskäyrä ja edistynyttä matematiikkaa sen luomiseksi. Se ei kuitenkaan ole uskottavaa, sillä käytössä ei ollut 1800-luvun havaintoja genetiikasta ja 1900-luvun havaintoja mutaatioista. Lisäksi Sanfordin kollegoineen tekemät simulaatiot tuottavat hyvin tarkasti samanlaisia käyriä.
”Mikä on genomin salaisuus? Sen pelkkä olemassaolo on sen salaisuus. Ihmisen ymmärryksen ylittävä informaatio ja monimutkaisuus on ohjelmoitu pölyhiukkastakin pienempään tilaan. Mutaatio ja valinta eivät voi alkuunkaan selittää sitä. On täysin selvää, ettei genomimme ole voinut syntyä itsestään. Ainoa perusteltu vaihtoehto itsestään syntyneelle genomille on suunniteltu genomi. Eikö tämä olekin pelottavan suuri salaisuus – tutkistelumme arvoinen?”
– Sanford62
Luku 11: Yhteenveto uusista tieteellisistä havainnoista
Sanford julkaisi kirjansa ensimmäisen painoksen vuonna 2005, ja tämän jälkeen on tehty monia uusia tieteellisiä havaintoja, jotka tukevat kirjan pääteesejä. Monet näistä on tehty simuloimalla Mendel’s accountant -ohjelmalla biologisia populaatioita. Ohjelma ei itsessään määrää, mikä lopputulos on, vaan se riippuu käytettävistä parametreista. Biologisesti realistisilla parametreilla populaatiot rappeutuvat ja kehitystä nähdään vain erittäin epärealistisilla parametreilla.
Sanfordin kollegojensa kanssa tuottama uusi kokeellinen todistusaineisto
Noin 90% haitallisista mutaatioista kertyy tasaisesti populaatioon, ja vain kaikkein pahimmat poistuvat. Lisäksi voidaan määrittää valintakynnys, jota vähäisemmät mutaatiot eivät tule valituksi. Kaikki muut biologiset ja geneettiset tekijät (kohina) nostavat valintakynnystä.63 Sama pätee myös hyödyllisten mutaatioiden valintaan, mutta näistä yli 98% jää realistisilla asetuksilla valinnan ulottumattomiin.64 Merkittävät hyödylliset mutaatiot tulevat kyllä valituiksi, mutta tämä samalla haittaa vähemmän merkittävien hyödyllisten mutaatioiden valintaa.64
Luvussa 7 esitetty mutaatioiden laskentamekanismi ei myöskään toimi realistisella todennäköisyysvalinnalla.50 Se voisi toimia vain, jos kaikkien mutaatioiden vaikutus olisi sama, mutta niin ei ole ja luonnonvalinnan pitäisi laskea kaikkien yksilöiden mutaatiot, ja sen jälkeen poistaa tarkasti kaikki ne, joilla on niitä eniten. Näin ei kuitenkaan todellisuudessa tapahdu.
Luvussa 7 esitettiin myös synergistisen epistasian mekanismi, joka ei toimi, vaikka simulaatioissa käytettäisiin evoluutioteorian kannalta hyvin suotuisia parametreja. Tällöinkin synergistinen epistasia vain kiihdyttää rappeutumista ja nopeuttaa sukupuuttoa.51
Avida-ohjelman on väitetty osoittavan, että evoluutioteoria on pätevä. Kyseinen ohjelma ei ole biologisten populaatioiden simulaatio-ohjelma, vaan eräänlainen tietokonepelisimulaatio. Avidassa jokainen hyödylliseksi merkitty muutos kaksinkertaistaa kelpoisuuden (eli lisääntymisnopeuden) ja jokainen haitallinen puolittaa sen. Näillä asetuksilla valinta on äärimmäisen tehokasta, mutta nämä rangaistukset ja palkinnot hyödyllisistä ja haitallisista mutaatioista ovat äärimmäisen rajusti liioiteltuja biologiseen todellisuuteen nähden. Mendel’s accountant-ohjelmalla saadaan samoilla lisääntymisvaikutuksilla aikaan samanlaista räjähtävää kehitystä, mutta jos hyödyllisten ja vahingollisten muutosten vaikutukset määritellään biologisesti realistisiksi, kehitystä ei tapahdukaan: hyödyllisiä muutoksia ei valita.65
Haldanen dilemma on vahvistettu. Walter ReMine havaitsi kehittämällään matemaattisella mallilla saman, mitä Haldane omissa analyyseissään.37 Simulaatiokokeilla on edelleen havaittu, että riippuen käytettävistä parametreista, 300 000 sukupolven aikana 10 000 yksilön populaatiossa voi valinta levittää populaatioon joistain sadoista joihinkin tuhansiin mutaatiota.55
Populaatiogenetiikassa on ollut ongelmana, että populaatioista tehdään liiaksi yksinkertaistettuja matemaattisia malleja, jotka eivät vastaa todellisuutta. Numeerisilla simulaatioilla voidaan kuitenkin realistisesti selvittää, miten mutaatio ja luonnonvalinta toimivat todellisuudessa. Tämän on havainnut Sanfordin lisäksi toinenkin numeerisia simulaatioita mutaatioiden ja valinnan analyysissään käyttävä tutkimusryhmä.66
Geneettisen entropian validointi todellisissa populaatioissa
Pitkäikäisillä lajeilla geneettisen entropian havaitseminen on vaikeaa, sillä geneettisen entropian vaikutukset tulevat näkyviin vasta useita sukupolvia myöhemmin. Tällaisilla lajeilla kokeet kestäisivät satoja vuosia. Bakteereillakin kokeet kestävät hyvin pitkään, koska mutaationopeus sukupolvea kohti on pientä. Kokeellisen näytön saamiseksi tarvittaisiin siis kohtuullisen suuri mutaationopeus ja lyhyt sukupolvi, ja nämä tekijät toteutuvat RNA-viruksilla.
Sanford on tutkinut H1N1-influenssavirusta, joka aiheutti suuren pandemian vuonna 1918.59 Ensin tehtiin numeerisia simulaatiokokeita influenssavirusta vastaavalla RNA-viruksmallilla.67 Näiden simulaatioiden tuloksena on, että haitallisia mutaatioita kertyy tasaisesti ja nopeasti viruksen perimään ja sen taudinaiheutuskyky laskee nopeasti. H1N1-virusnäytteitä on kerätty vuosikymmenten ajan, ja näistä näytteistä viruksen perimä on sekvensoitu, joten tätä aineistoa voitiin käyttää simulaatioiden tulosten vertailemiseksi todelliseen aineistoon. Myös tästä aineistosta havaittiin, että mutaatioita kertyi tasaisesti ja viruksen taudinaiheutuskyky laski samaa tahtia. Virus katosikin vuonna 2009, kun 10% viruksen perimästä oli mutatoitunut. Tämä vertailu vahvisti, että Mendel’s accountant -ohjelmalla saadaan simuloitua todellisuutta vastaavasti mutaatioiden ja valinnan toimintaa.
Sanford on myös arvioinut uudestaan Lenskin e.coli-bakteerikokeen.15, 16 Koe on jatkunut vuodesta 1988, mutta on silti vielä hyvin lyhyt yli 50 000 solunjakautumisesta huolimatta. Bakteereilla mutaationopeus on ainakin tuhat kertaa RNA-viruksia pienempi ja Lenskin e.coli-bakteereihin on kertynyt vain muutama sata mutaatiota yli neljän miljoonan nukleotidin perimään, samalla kun H1N1-viruksella yli 10% perimästä mutatoitui. Näin ollen RNA-viruksilla havaittuja rappeutumisen ääri-ilmiöitä bakteereilla nähdäkseen Lenskin kokeen pitäisi jatkua vuosisatoja.
Tarkemmin katsottuna Lenskin aineisto kuitenkin osoittaa rappeutumista. E.coli -bakteerit elävät hyvin keinotekoisessa ympäristössä, ja tähän ympäristöön sopeuttavat muutokset ovat aina hajottaneet jonkin toiminnon tai säätelymekanismin. Tässä keinotekoisessa ympäristössä bakteerit eivät tarvitse puoliakaan genominsa informaatiosta, joten tässä ympäristössä eläessään näiden turhien toimintojen ja säätelymekanismien tuhoutuminen jatkuu. Lopulta bakteerit ovat sopeutuneet niin, että ne voivat elää vain tässä keinotekoisessa ympäristössä, jolloin on saavutettu suurin energiatehokkuus ja näin tehokkain lisääntyminen juuri tässä ympäristössä. Tähän tilanteeseen pääsemiseen voi mennä vuosisatoja.
Joidenkin toiminnon hajottaneiden mutaatioiden on havaittu sopeuttaneen bakteereita ympäristöönsä, mutta suurimmalla osalla ei ole havaittu vaikutusta, joten näiden voidaan olettaa olevan lähes neutraaleja, hyvin hienoisesti vahingollisia muutoksia. Nämä ajavat bakteerit ennen pitkää sukupuuttoon.
Muut tutkimustulokset
DNA-koodin päällekkäisistä koodeista on tehty tutkimusta.14 Fisherin teoreema (Fundamental Theorem of Natural Selection) on osoitettu virheelliseksi – mutaatioiden kokonaisvaikutus ei ole neutraali, vaan negatiivinen.68
Biological Information – New Perspectives -kirja on julkaistu69, massiivinen ENCODE-projekti on osoittanut, että suurin osa ihmisen geeniperimästä on toiminnallista70, 2 ja myös Lynchin julkaisu vuodelta 2010 osoittaa ihmisen genomin rappeutuvan nopeasti.19
Liite 4: Voivatko geeniduplikaatio ja polyploidia kasvattaa geneettistä informaatiota?
Duplikaatiossa yhdestä tai muutamasta nukleotidista yhteen tai kaikkiin kromosomeihin voivat kopioitua toiseksi kopioksi. Polyploidiasta on kyse, kun kaikki kromosomit kahdentuvat. Voivatko nämä tuottaa uutta informaatiota?
Jos tämän artikkelin kaikki kirjaimet tai kaikki kappaleet olisi kirjoitettu kahteen kertaan, tekisikö se tästä artikkelista paremman? On selvää, että kaikki tällaiset kahdentumiset laajuudesta riippumatta ovat vahingollisia. Duplikaatioita uuden informaation luojana pitävät vain ne, jotka uskovat kaikkivoipaan valintaan, mutta on selvää, että siinä, missä valinta voi hidastaa perimän tuhoutumista, se ei voi pysäyttää sitä. Suurin osa duplikaatioistakin on vahingollisia ja lähes neutraaleita.
Jos ihmisen koko perimä kahdentuu, se johtaa kuolemaan. Jos yksi kromosomi kahdentuu, se johtaa kuolemaan, jos kyseessä on iso kromosomi, pienempien kromosomien tapauksessa se johtaa vakaviin ongelmiin, esimerkiksi 21-kromosomin ylimääräinen kopio Downin oireyhtymään. Tunnetaan myös lukuisia pienempiä duplikaatioita, jotka aiheuttavat perinnöllisiä sairauksia.
Jos johonkin geeniin on tullut vahingollisia muutoksia ja tämä sitten duplikoituu, ei tämä lisää informaatiota. Itse asiassa tämä heikentää valintaa, sillä tällöin geenille on olemassa varmuuskopio, joten vahingolliset muutokset eivät tule niin nopeasti esiin. Duplikaation jälkeen kahdentuneista geeneistä toinen ei myöskään pysy muuttumattomana, vaan siihenkin tulee mutaatioita yhtä nopeasti kuin toiseen kopioonkin. Näin ne rappeutuvat molemmat käytännössä yhtä nopeasti. Lisäksi geenien konversio voi aiheuttaa mutaatioiden siirtymistä kahdentuneiden geenien välillä, jolloin myös se geeni, johon mutaatiota ei alun perin tullut, saa sen.
Koska on olemassa polyploidisia kasveja, on väitetty, että nämä osoittavat duplikaatioiden olevan hyödyllisiä. Jos kasvien ei-sukulinjaan kuuluvia soluja altistetaan kolkkisiini-nimiselle kemikaalille, se saa aikaan solunjakautumiseen kromosomit kahdentavan häiriön. Tällaiset kasvit ovat lisääntymiskyvyttömiä ja kitukasvuisia, sillä ne tuhlaavat kaksi kertaa enemmän energiaa DNA:n tuottamiseen ilman, että tästä on mitään hyötyä kasville. Lisäksi iso määrä geenejä säätelevästä informaatiosta riippuu geenien määrästä, joten nämä säätelymekanismit rikkoutuvat ja näin menetetään informaatiota.
Jos polyploidia saa alkunsa sukusoluista (suvullinen polyploidia), joissa kumpikin on jäänyt vähennysjakautumatta, niin tällöin kyseisen yksilön informaatio kasvaa, sillä molempien vanhempien koko informaatio yhdistyy tällä yksilöllä. Kokonaisinformaatio ei kuitenkaan kasva, sillä kahden vanhemman vain jo olemassa ollut informaatio yhdistyy.
Suvullisessa polyploidiassa yhdellä kasvilla voi olla neljä versiota samasta geenistä (esim. peruna). Näin ollen kolme näistä voi mutatoitua rikki, mutta niin kauan kuin neljäs toimii, kasvilla ei ole mitään hätää. Näin ne toimivat varmuuskopioina hidastamassa rappeutumista. Hintana tällaisesta varmuuskopiojärjestelmästä on se, ettei valinta pysty poistamaan mutaatioita erityisen tehokkaasti, jolloin pitkällä aikavälillä rappeutuminen on vielä varmempaa.
Jos polyploidian hyötynä on lyhyellä aikavälillä rappeutumisen hidastaminen, sama pätee myös saman geenin eri puolilla genomia olevien kopioiden hyötyihin. Yleensä kun tällainen kopio löytyy, oletetaan vain sen olevan seurausta muinaisesta duplikaatiosta. Lisäksi yleensä oletetaan, että mutaatiot ovat myöhemmin tuottaneet eroja näihin kopioihin. Tämä on kuitenkin vain teoreettista päättelyä, eikä havaintoja. Saman geenin eri kopiot voivat selittyä myös varmuuskopioina tai monimutkaisen geenien säätelyn osana.
Duplikaatio voi joissain erittäin harvinaisissa tapauksissa olla itsessään hyödyllinen, tällöin kyse on geenin ilmentymän voimistumisesta. Se on lähes aina vahingollista, sillä yhden geenin ilmentymisen muutos häiritsee myös muiden geenien ilmentymistä. Tämä näkyy erityisesti geenimuuntelua toteutettaessa, jolloin kasveille tulee erilaisia geneettisiä vikoja satunnaisiin genomin paikkoihin lisätyistä geeneistä. Samalla tavalla duplikaatiotkin rikkovat genomin kokonaisfunktiota. Joskus harvoin kuitenkin geenin ilmentymisen voimistuminen voi olla hyödyllistä. Tähän tarkoitukseen duplikaatio on kuitenkin hyvin tehoton tapa, eikä kehitys voi olla aina näin johdonmukaisen tehotonta.
Ajatus siitä, että geeniduplikaation kautta voi syntyä uutta informaatiota, johtuu siitä, ettei evoluutiota voida nähdä muuten tapahtuneen. Lisäksi kun tätä ajatusta toistetaan riittävän kauan, siitä tulee totta. Todellisuutta tämä käsitys ei kuitenkaan vastaa.
Kirjallisuus
- Sanford, John C.: Eliömaailma rappeutuu – geneettinen entropia ja perimän salaisuus. Datakirjat, 2015. 978-952-68258-1-6.
Aiheesta muualla
- Dr John Sanford: Kriitikko ei välitä tosiasioista Genetic Entropy -kirjan arviossaan luominen.fi. Viitattu 4.7.2020.
- geneticentropy.org -verkkosivu
Viitteet
- ^ Sanford, s. 25
- > 2,0 2,1 The ENCODE Project Consortium: An integrated encyclopedia of DNA elements in the human genome. Nature, 2012, nro 489, s. 57-74.
- ^ Eyre-Walker, A. ja Keightley, P.D.: The distribution of fitness effects of new mutations. Nat Rev Genet, 2007, nro 8, s. 610-618.
- ^ Kimura M.: Model of effective neutral mutations in which selective constraint is incorporated. PNAS, 1979, nro 76, s. 3440-3444.
- ^ https://luominen.fi/css_images/kimuras_distribution_question.png
- > 6,0 6,1 6,2 Muller H.J.: The relation of recombination to mutational advance. Mutation research, 1964, nro 1, s. 2-9.
- ^ Sanford, s. 30
- ^ Gerrish P.J ja R. Lenski: The fate of competing benefical mutations in an asexual population. Genetica, 1998, nro 102/103, s. 127-144.
- ^ Bataillon T.: Estimaton of spontaneous genome-wide mutation rate parameters: whither benefical mutations?. Heredity, 2000, nro 84, s. 497-501.
- ^ Elena S.F. ym.: Distribution of fitness effects caused by random insertion mutations in bacteria. Genetica, 1998, nro 102/103, s. 349-358.
- ^ Sanford, s. 32
- ^ Sanford, s. 33
- ^ Bergman J.: Research on the deterioration of the genome and Darwinism: why mutations result in degeneration of the genome. (Intelligent design Conference, Biola University) ', 22.-23.4.2004.
- > 14,0 14,1 Montañez, G., Marks R., Fernandez J. ja Sanford J.: Multiple overlapping genetic codes profoundly reduce the probability of beneficial mutation. Marks II R.J. ym. (toim.) Biological Information – New Perspectives, 2013, s. 139-167. Artikkelin verkkoversio (viitattu 3.7.2020).
- > 15,0 15,1 Barrick, J.E. ym.: Genome evolution and adaptation in a long-term experiment with Escherichia coli. Nature, 2009, nro 461, s. 1243-1247.
- > 16,0 16,1 Lenski, R.E.: Evolution in Action: a 50,000-generation salute to Charles Darwin. Microbe, 2011, nro 6(1), s. 30-33.
- ^ Bataillon, T. ja Bailey, S.F.: Effects of new mutations on fitness: insights from models and data. Ann. N.Y. Acad. Sci., 2014, s. 1-17. Artikkelin verkkoversio (viitattu 3.7.2020).
- ^ Muller, H.J.: Our load of mutations. Amer. J. Human Genetics, 1950, nro 2, s. 111-176.
- > 19,0 19,1 19,2 Lynch, M.: Rate, molecular spectrum, and consequences of human mutation. PNAS, 2010, nro 107 (3), s. 961-968.
- ^ Nachman, M.W. ja Crowell, S.L.: Estimate of the mutation rate per nucleotide in humans. Genetics, 2000, nro 156, s. 297-304.
- > 21,0 21,1 Kondrashov, A.S.: Direct Estimate of human per nucleotide mutation rates at 20 loci causing Mendelian diseases. Human Mutation, 2002, nro 21, s. 12-27.
- ^ Campbell, C.D. ja Eichler. E.E.: Properties and rates of germline mutations in humans. Trends in Genetics, 2013, nro 29, s. 575-584.
- ^ Xue, Y. ym.: Human Y chromosome base-substitution mutation rate measured by direct sequencing in a deep-rooting pedigree. Current Biology, 2009, nro 19, s. 1453-1457.
- ^ Sanford, s. 36
- ^ Parsons, T.J. ym.: A high observed substitution rate in the human mitochondrial DNA control region. Nature Genetics, 1997, nro 15, s. 363-368.
- ^ Sutherland, G.R. ja R.I. Richards: Simple tandem repeats and human disease. PNAS, 1995, nro 92, s. 3636-3641.
- ^ Ellegren, H.: Microsatellite mutations in the germline: implications for evolutionary inference. TIG, 2000, nro 16, s. 551-558.
- ^ Sanford, s. 42-43
- ^ Bernardes, A.T.: Mutation load and the extinction of large populations. Physica ACTA, 1996, nro 230, s. 156-173.
- ^ Crow, J.F.: The high spontaneous mutation rate: is it a health risk?. PNAS, 1997, nro 94, s. 8380-8386.
- ^ Sanford, s. 50
- ^ Sanford, s. 50-51
- > 33,0 33,1 33,2 33,3 Haldane, J.B.S.: The cost of natural selection.. J. Genetics, 1957, nro 55, s. 511-524.
- > 34,0 34,1 34,2 Kimura, M. ja Ohta, T.: Theoretical Aspects of Population Genetics. Princeton University Press, Princeton, NJ, 1971, s. ss. 26-31, s. 53.
- > 35,0 35,1 35,2 35,3 Kimura, M.: Neutral Theory of Molecular Evolution, s. s. 26, ss. 30-31. Cambridge Univ. Press, 1983.
- > 36,0 36,1 ReMine, W.: The Biotic Message: Evolution versus Message Theory. St. Paul Science, St. Paul, MN, 1993.
- > 37,0 37,1 37,2 ReMine, W.: Cost of Selection Theory. Technical Journal, 2005, nro 19, s. 113-125.
- > 38,0 38,1 Kimura, M.: Evolutionary rate at the molecular level. Nature, 1968, nro 217, s. 624-626.
- ^ Ohta, T.: Slightly deleterious mutant substitutions in evolution. Nature, 1973, nro 246, s. 96-98.
- ^ Ohta, T.: Mutational pressure as the main cause of molecular evolution and polymorphism. Nature, 1974, nro 252, s. 351-354.
- ^ Ohta, T.: The nearly neutral theory of molecular evolution. Ann Rev Ecol Syst, 1992, nro 23, s. 263-286.
- ^ Ohta, T.: Near-neutrality in evolution of genes and gene regulation. PNAS, 2002, nro 99, s. 16134-16137.
- > 43,0 43,1 43,2 Kondrashov, A.S.: Contamination of the genome by very slightly deleterious mutations: why have we not died 100 times over?. J. Theor. Biol., 1995, nro 175, s. 583-594.
- ^ Sanford, s. 57
- ^ Lynch, M., Conery J., ja Burger R.: Mutation accumulation and the extinction of small populations. Am. Nat., 1995, nro 146, s. 489-518.
- > 46,0 46,1 46,2 Tishkoff, S.A. ja Verrelli, B.C.: Patterns of human genetic diversity: implications for human evolutionary history and disease.. Annual Review of Genomics and Human Genetics, 2003, nro 4, s. 293-340.
- > 47,0 47,1 Loewe, L.: Quantifying the genome decay paradox due to Muller’s ratchet in human mitochondrial DNA. Genetic Research, 2006, nro 87, s. 133-159.
- ^ Merila J. ja B.C. Shelton: Lifetime reproductive success and heritability in nature. The American Naturalist, 2000, nro 155, s. 301-310.
- ^ Kruuk, L.E.B. ym.: Heritability of fitness in a wild mammal population. PNAS, 2000, nro 97, s. 698–703.
- > 50,0 50,1 Brewer, W., J. Baumgardner ja J. Sanford: Using Numerical Simulation to Test the “Mutation-Count” Hypothesis. Marks II R.J. ym. (toim.) Biological Information – New Perspectives, 2013, s. 298-311. [www.worldscientific.com/doi/pdf/10.1142/9789814508728_0012 Artikkelin verkkoversio] (viitattu 4.7.2020).
- > 51,0 51,1 Baumgardner J., W. Brewer ja J. Sanford: Can Synergistic Epistasis Halt Mutation Accumulation? Results from Numerical Simulation. Marks II R.J. ym. (toim.) Biological Information – New Perspectives, 2013, s. 312-337. [www.worldscientific.com/doi/pdf/10.1142/9789814508728_0013 Artikkelin verkkoversio] (viitattu 4.7.2020).
- ^ Felsenstein, J.: The evolutionary advantage of recombination. Genetics, 1974, nro 78, s. 737-756.
- ^ Patterson, C.: Evolution. Comstock Publishing Associates, Ithaca, NY, 1999.
- ^ Crow, J.F. and M. Kimura: An Introduction to Population Genetics Theory, s. 249. Harper and Row. NY, NY, 1970.
- > 55,0 55,1 Rupe, C.L. ja J.C. Sanford.: Using numerical simulation to better understand fixation rates, and establishment of a new principle: Haldane’s Ratchet. ICC, 2013. Artikkelin verkkoversio.
- > 56,0 56,1 Gabriel, S.B. ym.: The structure of haplotype blocks in the human genome. Science, 2002, nro 296, s. 2225-2229.
- ^ Yelin, R. ym.: Widespread occurrence of antisense transcription in the human genome. Nature Biotechnology, 2003, nro 21, s. 379-386.
- ^ Britten, R.J.: Divergence between samples of chimpanzee and human DNA sequences is 5% counting indels. PNAS, 2002, nro 99, s. 13633-13635.
- > 59,0 59,1 Carter, R.W. ja J.C. Sanford: A new look at an old virus: patterns of mutation accumulation in the human H1N1 influenza virus since 1918. Theoretical Biology and Medical Modeling, 2012, nro 9, s. 42. doi:10.1186/1742-4682-9-42. Artikkelin verkkoversio.
- ^ Simonsen L. ym.: Pandemic versus epidemic mortality: a pattern of changing age distribution. Journal of Infectious Diseases, 1998, nro 178, s. 53-60.
- ^ Holladay, P.M. ja J.M. Watt: De-generation: an exponential decay curve in old testament genealogies. (52 nd Natl. Conf., Nashville, TN Nov. 15-17, 2000) Evangelical Theological Society Papers, 2001, nro 2001.
- ^ Sanford,s. 116
- ^ Gibson, P., J. Baumgardner, W. Brewer, ja J. Sanford: Can Biological Information Be Sustained By Purifying Natural Selection?. Marks II R.J. ym. (toim.) Biological Information – New Perspectives, 2013, s. 232-263. Artikkelin verkkoversio (viitattu 12.7.2020).
- > 64,0 64,1 Sanford, J., J. Baumgardner, ja W. Brewer: Selection Threshold Severely Constrains Capture of Beneficial Mutations. Marks II R.J. ym. (toim.) Biological Information – New Perspectives, 2013, s. 264-297. Artikkelin verkkoversio (viitattu 12.7.2020).
- ^ Nelson, C.W. and J.C. Sanford: The Effects of Low-Impact Mutations in Digital Organisms. Theoretical Biology and Medical Modeling Vol. 8, huhtikuu 2011, s. 9. Artikkelin verkkoversio.
- ^ Qui, S., A. McSweeny, S. Choulet, A. Saha-Mandal, L. Fedorova, ja A. Fedorov: Genome evolution by matrix algorithms (GEMA): cellular automata approach by population genetics. Genome Biol. Evol., 2014, nro 6(4), s. 988-999.
- ^ Brewer, W., F. Smith and J. Sanford: Information loss: potential for accelerating natural genetic attenuation of RNA viruses. Marks II R.J. ym. (toim.) Biological Information – New Perspectives, 2013, s. 369-384. Artikkelin verkkoversio (viitattu 12.7.2020).
- ^ Basener, William F. Ja John C. Sanford: The fundamental theorem of natural selection with mutations. Journal of Mathematical Biology, 2018, nro 76, s. 1589–1622. Artikkelin verkkoversio (viitattu 13.7.2020).
- ^ Marks, II R.J., M.J. Behe, W.A. Dembski, B.L. Gordon, ja J.C. Sanford: Biological Information – New Perspectives, s. 1-559. World Scientific Publishing Co., Singapore, 2013. Teoksen verkkoversio.
- ^ The ENCODE Project Consortium: Identification and analysis of functional elements in 1% of the human genome by the ENCODE pilot project. Nature, 2007, nro 447, s. 799-816.
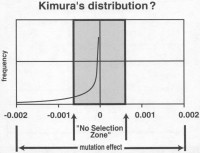{kind=link}